Het actieve centrum van het eiwit en zijn interactie met het ligand. Eiwitfunctie
Het actieve centrum van eiwitten is een bepaald deel van een eiwitmolecuul, in de regel gelokaliseerd in zijn depressie, gevormd door radicalen van aminozuren verzameld op een bepaalde ruimtelijke plaats tijdens de vorming van een tertiaire structuur en in staat om complementair te binden aan een ligand . In de lineaire volgorde van de polypeptideketen kunnen de radicalen die het actieve centrum vormen zich op aanzienlijke afstand van elkaar bevinden.
De hoge specificiteit van binding van het eiwit aan het ligand wordt geleverd door de complementariteit van de structuur van de actieve plaats van het eiwit en de structuur van het ligand.
Complementariteit wordt opgevat als de ruimtelijke en chemische overeenkomst van op elkaar inwerkende moleculen. Het ligand moet de conformatie van de actieve plaats kunnen binnendringen en ruimtelijk samenvallen. Dit samenvallen kan onvolledig zijn, maar vanwege de conformationele labiliteit van het eiwit is het actieve centrum in staat tot kleine veranderingen en wordt het "aangepast" aan het ligand. Bovendien moeten er bindingen ontstaan tussen de functionele groepen van het ligand en de aminozuurradicalen die de actieve plaats vormen, die de ligand op de actieve plaats houden. De bindingen tussen het ligand en het actieve centrum van het eiwit kunnen zowel niet-covalent (ionisch, waterstof, hydrofoob) als covalent zijn.
Kenmerken van het actieve centrum
Het actieve centrum van een eiwit is een gebied dat relatief geïsoleerd is van de omringende eiwitomgeving, gevormd door aminozuurresiduen. In dit gebied vormt elk residu, vanwege zijn individuele grootte en functionele groepen, het "reliëf" van het actieve centrum.
Unieke eigenschappen actieve centrum zijn niet alleen afhankelijk van de chemische eigenschappen van de aminozuren die het vormen, maar ook van hun exacte onderlinge oriëntatie in de ruimte. Daarom zelfs kleine schendingen van de algemene conformatie van het eiwit als gevolg van puntveranderingen in de primaire structuur of omstandigheden omgeving kan leiden tot een verandering in de chemische en functionele eigenschappen van de radicalen die het actieve centrum vormen, de binding van het eiwit aan het ligand en zijn functie verstoren. Tijdens denaturatie wordt het actieve centrum van eiwitten vernietigd en gaat hun biologische activiteit verloren.
Het actieve centrum is vaak zo gevormd dat de toegang van water tot de functionele groepen van zijn radicalen beperkt is, d.w.z. omstandigheden worden gecreëerd voor het ligand om te binden aan aminozuurradicalen.
De ligand-eiwitbindingsplaats bevindt zich vaak tussen domeinen. Het proteolytische enzym trypsine, dat betrokken is bij de hydrolyse van peptidebindingen van voedingseiwitten in de darm, heeft bijvoorbeeld 2 domeinen die door een groef worden gescheiden. Het binnenoppervlak van de groef wordt gevormd door de aminozuurradicalen van deze domeinen, die ver van elkaar in de polypeptideketen liggen (Ser 177, His 40, Asp 85).
Verschillende domeinen in een eiwit kunnen bij interactie met een ligand ten opzichte van elkaar bewegen, wat de verdere werking van het eiwit vergemakkelijkt. De belangrijkste eigenschap van eiwitten die aan hun functies ten grondslag liggen, is de selectiviteit van de hechting van specifieke liganden aan bepaalde delen van het eiwitmolecuul.
Verscheidenheid aan liganden:
Liganden kunnen anorganische (vaak metaalionen) en organische stoffen zijn, stoffen met een laag en hoog molecuulgewicht;
Er zijn liganden die hun chemische structuur veranderen wanneer ze zich hechten aan de actieve plaats van het eiwit (veranderingen in het substraat in de actieve plaats van het enzym);
Er zijn liganden die zich pas aan een eiwit binden op het moment van functioneren (bijvoorbeeld O 2 getransporteerd door hemoglobine), en liganden die permanent aan een eiwit gebonden zijn die een hulprol spelen bij het functioneren van eiwitten (bijvoorbeeld ijzer, dat deel uitmaakt van hemoglobine).
HOOFDSTUK 3
BOEREN. MECHANISME VAN ENZYM ACTIE
Enzymen of enzymen zijn specifieke eiwitten die deel uitmaken van alle cellen en weefsels van levende organismen en de rol spelen van biologische katalysatoren.
Algemene eigenschappen van enzymen en anorganische katalysatoren:
1. Niet verbruikt tijdens de reactie.
2. Oefen hun effect uit bij lage concentraties.
3. Beïnvloed de waarde van de reactie-evenwichtsconstante niet.
4. Hun actie is onderworpen aan de wet van de massa's op het werk.
5. Versnel thermodynamisch onmogelijke reacties niet.
Verschillen tussen enzymen en anorganische katalysatoren.
1. Thermolabiliteit van enzymen.
2. Afhankelijkheid van enzymactiviteit van de pH van het medium.
3. Specificiteit van enzymwerking.
4. De snelheid van enzymatische reacties is onderhevig aan bepaalde kinetische wetten.
5. De activiteit van enzymen hangt af van de werking van regulatoren - activatoren en remmers.
6. Een aantal enzymen ondergaan tijdens de vorming van de tertiaire en quaternaire structuren een postsynthetische modificatie.
7. De grootte van het enzymmolecuul is meestal veel groter dan de grootte van hun substraten.
Enzym molecuul structuur
In structuur kunnen enzymen eenvoudige of complexe eiwitten zijn. Een enzym dat een complex eiwit is, wordt holo-enzym... Het eiwitgedeelte van het enzym wordt het apoenzym genoemd, het niet-eiwitgedeelte wordt de cofactor genoemd. Onderscheiden twee soorten cofactoren:
1. Prothetische groep - stevig geassocieerd met het apo-enzym, vaak covalente bindingen.
2. Co-enzym is een niet-eiwitdeel dat gemakkelijk kan worden gescheiden van het apo-enzym. Derivaten van vitamines worden vaak gebruikt als co-enzymen.
naar co-enzymen omvatten de volgende verbindingen:
Derivaten van vitamines;
Hemes die deel uitmaken van cytochromen, catalase, peroxidase, guanylaatcyclase, NO-synthase en een prosthetische groep van enzymen zijn;
Nucleotiden - donoren en acceptoren van het fosforzuurresidu;
Ubiquinon of co-enzym Q, dat betrokken is bij de overdracht van elektronen en protonen in de weefselademhalingsketen;
Fosfoadenosylfosfosulfaat, dat betrokken is bij de overdracht van sulfaat;
Glutathion, dat betrokken is bij redoxreacties.
Tabel 3.1.
Co-enzymfuncties van vitamines
Ppt% 5C34928-slozhnye_belki_ch1_1.jpg "alt =" (! TAAL:> Het actieve centrum van het eiwit en zijn interactie met het ligand. Tijdens de vorming van de tertiaire structuur"> Активный центр белка и его взаимодействие с лигандом. В процессе формирования третичной структуры на поверхности функционально активного белка, обычно в углублении, образуется участок, сформированный радикалами аминокислот, далеко стоящими друг от друга в первичной структуре. Этот участок, имеющий уникальное строение для данного белка и способный специфично взаимодействовать с определенной молекулой или группой похожих молекул, называется центром связывания белка с лигандом или активным центром. Лигандами называются молекулы, взаимодействующие с белками.!}
Ppt% 5C34928-slozhnye_belki_ch1_2.jpg "alt =" (! LANG:> Een ligand kan een stof met een laag molecuulgewicht of een stof met een hoog molecuulgewicht (macromolecuul) zijn, inclusief"> Лигандом может быть как низкомолекулярное, так и высокомолекулярное (макромолекула) вещество, в том числе и другой белок. Лигандами являются субстраты ферментов, кофакторы, ингибиторы и активаторы ферментов, протомеры в олигомерном белке и т.д.!}
Ppt% 5C34928-slozhnye_belki_ch1_3.jpg "alt =" (! TAAL:> De hoge specificiteit van de interactie van het eiwit met het ligand wordt geleverd door de complementariteit van de structuur van het actieve centrum met de structuur van het ligand.">!}
Ppt% 5C34928-slozhnye_belki_ch1_4.jpg "alt =" (! TAAL:> Complementariteit is de ruimtelijke en chemische overeenkomst van op elkaar inwerkende oppervlakken. Een actieve site moet niet alleen"> Комплементарность - это пространственное и химическое соответствие взаимодействующих поверхностей. Активный центр должен не только пространственно соответствовать входящему в него лиганду, но и между функциональными группами радикалов, входящих в активный центр, и лигандом должны образоваться связи чаще всего нековалентные (ионные, водородные, а также гидрофобные взаимодействия), которые удерживают лиганд в активном центре.!}
Ppt% 5C34928-slozhnye_belki_ch1_5.jpg "alt =" (! LANG:> Complementaire eiwit-ligand interactie">!}
Ppt% 5C34928-slozhnye_belki_ch1_6.jpg "alt ="> ">
Ppt% 5C34928-slozhnye_belki_ch1_7.jpg "alt ="> ">
Ppt% 5C34928-slozhnye_belki_ch1_8.jpg "alt =" (! TAAL:> EIWITCLASSIFICATIE 1. Eenvoudige eiwitten bestaan alleen uit aminozuren. 2. Complexe eiwitten (holoproteïnen)"> КЛАССИФИКАЦИЯ БЕЛКОВ 1. Простые белки состоят только из аминокислот. 2. Сложные белки (холопротеины) содержат белковую часть (апопротеин) и небелковую (простетическую) группу.!}
Ppt% 5C34928-slozhnye_belki_ch1_9.jpg "alt =" (! TAAL:> Verschillende organische (lipiden, koolhydraten) en anorganische (metalen) stoffen kunnen fungeren als een prothetische groep."> В качестве простетической группы могут выступать различные органические (липиды, углеводы) и неорганические (металлы) вещества. Связь между простетической группой и апопротеином может быть как ковалентная, так и нековалентная. Простетическую группу порой можно рассматривать в качестве лиганда. Наличие небелковой части обеспечивает выполнение белком его функции. При утрате простетической группы холопротеин теряет свою активность.!}
Ppt% 5C34928-slozhnye_belki_ch1_10.jpg "alt =" (! TAAL:> Complexe eiwitten - chromoproteïnen - nucleoproteïnen - lipoproteïnen - fosfoproteïnen - glycoproteïnen - metalloproteïnen">!}
Ppt% 5C34928-slozhnye_belki_ch1_11.jpg "alt =" (! TAAL:> Metalloproteïnen omvatten holo-enzymen die niet-heem-gecoördineerde metaalionen bevatten. Onder de metalloproteïnen bevinden zich eiwitten,"> Металлопротеинам можно отнести холоферменты, содержащие негемовые координационно связанные ионы металлов. Среди металлопротеинов есть белки, выполняющие депонирующие и транспортные функции (например, железосодержащие ферритин и трансферрин) и ферменты (например, цинксодержащая карбоангидраза и различные супероксиддисмутазы, содержащие в качестве активных центров ионы меди, марганца, железа и других металлов). Но и хромопротеины, содержащие ионы металлов, также можно отнести к металлопротеинам.!}
Ppt% 5C34928-slozhnye_belki_ch1_12.jpg "alt =" (! TAAL:> Metalloproteïnen zijn vaak enzymen. Metaalionen in dit geval: - nemen deel aan de oriëntatie van het substraat"> Металлопротеины часто являются ферментами. Ионы металлов в этом случае: - участвуют в ориентации субстрата в активном центре фермента, входят в состав активного центра фермента и участвуют в катализе, являясь, например, акцепторами электронов на определенной стадии ферментативной реакции. Часто ион металла в составе фермента называют кофактором.!}
Ppt% 5C34928-slozhnye_belki_ch1_13.jpg "alt =" (! LANG:> Enzymatische metalloproteïnen omvatten eiwitten die, bijvoorbeeld: - koper - cytochroomoxidase, in een complex"> К ферментативным металлопротеинам относятся белки, содержащие например: - медь – цитохромоксидаза, в комплексе с другими ферментами дыхательной цепи митохондрий участвует в синтезе АТФ, - железо – ферритин, депонирующий железо в клетке, трансферрин, переносящий железо в крови, каталаза, обезвреживающая перекись водорода, - цинк – алкогольдегидрогеназа, обеспечивающая метаболизм этанола и других спиртов, лактатдегидрогеназа, участвующая в метаболизме молочной кислоты, - карбоангидраза, образующая угольную кислоту из CO2 и H2O, - щелочная фосфатаза, гидролизующая фосфорные эфиры !} verschillende verbindingen, - α2-macroglobuline, antiprotease-eiwit in het bloed. - selenium - schildklierperoxidase, betrokken bij de synthese van schildklierhormonen, het antioxidant-enzym glutathionperoxidase, - calcium - α-amylase van speeksel en pancreassap, hydrolyserend zetmeel.
Ppt% 5C34928-slozhnye_belki_ch1_14.jpg "alt =" (! TAAL:> Ferritine">!}
Ppt% 5C34928-slozhnye_belki_ch1_15.jpg "alt =" (! TAAL:> Fosfoproteïnen zijn eiwitten waarin een fosfaatgroep aanwezig is. Het bindt aan de peptideketen"> Фосфопротеины – это белки, в которых присутствует фосфатная группа. Она связывается с пептидной цепью через остатки тирозина, серина и треонина, т.е. тех аминокислот, которые содержат ОН-группу. Способ присоединения фосфата к белку на примере серина и тирозина!}
Ppt% 5C34928-slozhnye_belki_ch1_16.jpg "alt =" (! TAAL:> Fosforzuur kan vervullen: - Een structurele rol, het verlenen van lading, oplosbaarheid en veranderende eigenschappen"> Фосфорная кислота может выполнять: - Структурную роль, придавая заряд, растворимость и изменяя свойства белка, например, в казеине молока, яичном альбумине. Наличие остатков фосфорной кислоты способствует связыванию кальция, что необходимо для формирования, например, костной ткани. - Функциональную роль. В клетке присутствует много белков, которые связаны с фосфатом не постоянно, а в зависимости от активности метаболизма. Белок может многократно переходить в фосфорилированную или в дефосфорилированную форму, что играет регулирующую роль в его работе.!}
Ppt% 5C34928-slozhnye_belki_ch1_17.jpg "alt =" (! LANG:> Fosforylering is het proces waarbij het fosforzuurresidu van het fosforylerende donoragens naar het substraat wordt overgebracht, gewoonlijk"> Фосфорилирование - процесс переноса остатка фосфорной кислоты от фосфорилирующего агента-донора к субстрату, как правило, катализируемый ферментами (киназами) и ведущий к образованию эфиров фосфорной кислоты. Дефосфорилирование (утрату остатка фосфорной кислоты) катализируют фосфатазы. АТФ + R-OH → АДФ + R-OPO3H2 R-OPO3H2 + Н2О → R-OH + Н3РО4!}
Ppt% 5C34928-slozhnye_belki_ch1_18.jpg "alt =" (! TAAL:> Voorbeelden: 1) glycogeensynthase en glycogeenfosforylase-enzymen 2) gefosforyleerde histonen binden minder sterk"> Примеры: 1) ферменты гликогенсинтаза и гликогенфосфорилаза 2) гистоны в фосфорилированном состоянии менее прочно связываются с ДНК и активность генома возрастает. Изменение конформации белка в фосфорилированном и дефосфорилированном состоянии!}
Ppt% 5C34928-slozhnye_belki_ch1_19.jpg "alt =" (! LANG:> Lipoproteïnen bevatten niet-covalent gebonden lipiden als prothetisch onderdeel. Lipiden, in het bijzonder"> Липопротеины содержат в качестве простетической части нековалентно связанные липиды. Липиды, в частности жиры, холестерол и его эфиры не растворяются в водных фазах организма, поэтому транспорт их кровью и лимфой осуществляется в виде комплексов с белками и фосфолипидами, которые называются липопротеинами.!}
Ppt% 5C34928-slozhnye_belki_ch1_20.jpg "alt =" (! TAAL:> Alle lipoproteïnen hebben een vergelijkbare structuur: de kern bestaat uit hydrofobe moleculen: triacylglycerolen, cholesterolesters en"> Все липопротеины имеют сходное строение: ядро состоит из гидрофобных молекул: триацилглицеролов, эфиров холестерола, а на поверхности находится монослой фосфолипидов, полярные группы которых обращены к воде, а гидрофобные погружены в гидрофобное ядро липопротеина. Кроме фосфолипидов, на поверхности находятся белки – аполипопротеины (апобелками). Их выделяют несколько видов: А, В, С, D. В каждом типе липопротеинов преобладают соответствующие ему апобелки. Аполипопротеины выполняют различные функции. Интегральные аполипопротеины являются структурными компонентами. Периферические аполипопротеины в плазме крови могут передаваться от одного типа липопротеинов к другим, определяя их дальнейшие превращения.!}
Ppt% 5C34928-slozhnye_belki_ch1_21.jpg "alt =" (! LANG:> Schema van de structuur van lipoproteïne De structuur van lipoproteïne">!}
Ppt% 5C34928-slozhnye_belki_ch1_22.jpg "alt =" (! LANG:> De structuur van lipoproteïnen in bloedplasma">!}
Ppt% 5C34928-slozhnye_belki_ch1_23.jpg "alt =" (! LANG:> Er zijn vier hoofdklassen van lipoproteïnen: - lipoproteïnen met hoge dichtheid (HDL), - lipoproteïnen met lage dichtheid (LDL),"> Выделяют четыре основных класса липопротеинов: -липопротеины высокой плотности (ЛПВП), -липопротеины низкой плотности (ЛПНП), -липопротеины очень низкой плотности (ЛПОНП), -хиломикроны (ХМ). Каждый из типов ЛП образуется в !} verschillende stoffen en transporteert bepaalde lipiden. De concentratie en verhouding van bepaalde lipoproteïnen in het bloed spelen een hoofdrol bij het ontstaan van een dergelijke algemene vasculaire pathologie als atherosclerose. HDL is antiatherogeen, LDL en VLDL zijn atherogeen.
Ppt% 5C34928-slozhnye_belki_ch1_24.jpg "alt ="> ">
Ppt% 5C34928-slozhnye_belki_ch1_25.jpg "alt =" (! TAAL:> Glycoproteïnen of glycoconjugaten zijn eiwitten die een koolhydraatcomponent bevatten die covalent aan een polypeptidebasis is gehecht."> Гликопротеины или, гликоконъюгаты – это белки, содержащие углеводный компонент, ковалентно присоединенный к полипептидной основе. Содержание углеводов в них варьирует от 1% до 98% по массе. Два подкласса белков, содержащих углеводы: ■ протеогликаны ■ гликопротеины!}
Beschrijving = "">
Ppt% 5C34928-slozhnye_belki_ch1_27.jpg "alt =" (! TAAL:> Glycoproteïnen worden gekenmerkt door een laag gehalte aan koolhydraten, die zijn gehecht: - door een N-glycosidische binding aan de NH2-groep van sommige"> Для гликопротеинов характерно невысокое содержание углеводов, которые присоединены: - N-гликозидной связью к NН2-группе какого-нибудь аминокислотного остатка, например, аспарагина; - О-гликозидной связью к гидроксильной группе остатка серина, треонина,тирозина, гидроксилизина.!}
Ppt% 5C34928-slozhnye_belki_ch1_28.jpg "alt =" (! TAAL:> Vorming van O- en N-glycosidebindingen in glycoproteïnen. 1 - N-glycosidebinding tussen de amidegroep"> Образование О- и N-гликозидных связей в гликопротеинах. 1 - N-гликозидная связь между амидной группой аспарагина и ОН-группой моносахарида; 2 - О-гликозидная связь между ОН-группой серина и ОН-группой моносахарида.!}
Ppt% 5C34928-slozhnye_belki_ch1_29.jpg "alt =" (! LANG:> Methode voor het koppelen van koolhydraten aan eiwitten">!}
Ppt% 5C34928-slozhnye_belki_ch1_30.jpg "alt =" (! TAAL:> De functies van glycoproteïnen zijn: 1. Structureel - bacteriële celwand, botmatrix, bijv. collageen, elastine."> Функцией гликопротеинов являются: 1. Структурная – клеточная стенка бактерий, костный матрикс, например, коллаген, эластин. 2. Защитная – например, антитела, интерферон, факторы свертывания крови (протромбин, фибриноген). 3. Рецепторная – присоединение эффектора приводит к изменению конформации белка-рецептора, что вызывает внутриклеточный ответ. 4. Гормональная – гонадотропный, адренокортикотропный и тиреотропный гормоны. 5. Ферментативная – холинэстераза, нуклеаза. 6. Транспортная – перенос веществ в крови и через мембраны, например, трансферрин, транскортин, альбумин, Na+,К+-АТФаза.!}
Ppt% 5C34928-slozhnye_belki_ch1_31.jpg "alt =" (! LANG:> Schema van de structuur van de eiwitreceptor">!}
Ppt% 5C34928-slozhnye_belki_ch1_32.jpg "alt =" (! TAAL:> Chromoproteïnen zijn de verzamelnaam voor complexe eiwitten met gekleurde prosthetische groepen van verschillende chemische aard."> Хромопротеины - собирательное название сложных белков с окрашенными простетическими группами различной химической природы. гемопротеины (содержат гем), ретинальпротеины (содержат витамин А), флавопротеины (содержат витамин В2), кобамидпротеины (содержат витамин В12).!}
Ppt% 5C34928-slozhnye_belki_ch1_33.jpg "alt =" (! TAAL:> Flavoproteïnen zijn enzymen van redoxreacties. Ze bevatten derivaten van vitamine B2 flavine mononucleotide (FMN) en flavine adenine dinucleotide"> Флавопротеины - это ферменты окислительно-восстановительных реакций. Содержат производные витамина В2 флавинмононуклеотид (ФМН) и флавинадениндинуклеотид (ФАД). Связываются данные простетические группы ковалентно и придают желтое окрашивание. Эти простетические группы являются производными изоаллоксазина.!}
Ppt% 5C34928-slozhnye_belki_ch1_34.jpg "alt =" (! LANG:> Isoalloxazine is een heterocyclische verbinding afgeleid van pteridine. Het isoalloxazinemolecuul bestaat uit drie aromatische ringen -"> Изоаллоксазин - гетероциклическое соединения, производное птеридина. Молекула изоаллоксазина состоит из трех ароматических колец - бензольного, пиримидинового, пиразинового.!}
Ppt% 5C34928-slozhnye_belki_ch1_35.jpg "alt =" (! TAAL:> Heme-bevattende chromoproteïnen. Structureel vergelijkbare ijzer- of magnesiumporfyrinen zijn opgenomen als een niet-eiwitcomponent."> Гемопротеины - гем-содержащие хромопротеины. В качестве небелкового компонента включают структурно сходные железо- или магнийпорфирины. Белковый компонент может быть разнообразным как по составу, так и по структуре. Основу структуры простетической группы большинства гемосодержащих белков составляет порфириновое кольцо, являющееся в свою очередь производным тетрапиррольного соединения – порфирина. Порфирин!}
Ppt% 5C34928-slozhnye_belki_ch1_36.jpg "alt =" (! LANG:> De porfyrinering is in staat coördinatieverbindingen te vormen met verschillende metaalionen."> Порфириновое кольцо способно образовывать координационные соединения с различными ионами металлов. В результате комплексообразования формируются металлопорфирины: содержащие ионы железа – гемоглобины, миоглобин, цитохромы, пероксидаза, каталаза и др. (красное окрашивание), содержщие ионы магния – хлорофилл (зеленое окрашивание). Витамин В12 (кобалимин) содержит координированный ион кобальта Со2+ в порфириноподобном макроцикле – коррине, состоящем из четырех частично гидрированных пиррольных колец (розовое окрашивание).!}
Ppt% 5C34928-slozhnye_belki_ch1_37.jpg "alt =" (! TAAL:> Chlorofyl b. Chlorofylen zijn betrokken bij fotosynthese.">!}
Ppt% 5C34928-slozhnye_belki_ch1_38.jpg "alt =" (! TAAL:> Cytochromen verschillen in de aminozuursamenstelling van peptideketens, het aantal ketens en zijn onderverdeeld in typen a, b,"> Цитохромы различаются аминокислотным составом пептидных цепей, числом цепей и разделяются на типы а, b, с, d. Цитохромы находятся в составе дыхательной цепи и цепи микросомального окисления. Степень окисления железа в составе цитохромов меняется в отличие от гемоглобина и миоглобина Fe2+ ↔ Fe3+!}
Ppt% 5C34928-slozhnye_belki_ch1_39.jpg "alt =" (! LANG:> Myoglobine (MB) is een eiwit dat voorkomt in rode spieren en waarvan de belangrijkste functie het opslaan"> Миоглобин (Мв) - белок, находящийся в красных мышцах, основная функция которого - создание запасов О2, необходимых при интенсивной мышечной работе. Мв - сложный белок, содержащий белковую часть - апоМв и небелковую часть - гем. Первичная структура апоМв определяет его компактную глобулярную конформацию и структуру активного центра, к которому присоединяется небелковая часть миоглобина - гем. Кислород, поступающий из крови в мышцы, связывается с Fe2+ гема в составе миоглобина. Мв - мономерный белок, имеющий очень высокое сродство к О2, поэтому отдача кислорода миоглобином происходит только при интенсивной мышечной работе, когда парциальное давление O2 резко снижается. Формирование пространственных структур и функционирование миоглобина.!}
Ppt% 5C34928-slozhnye_belki_ch1_40.jpg "alt =" (! LANG:> Vorming van de MB-conformatie."> Формирование конформации Мв. В красных мышцах на рибосомах в ходе трансляции идет синтез первичной структуры Мв, представленной специфической последовательностью 153 аминокислотных остатков. Вторичная структура Мв содержит восемь α-спиралей, называемых !} met Latijnse letters van A tot H, waartussen zich niet-spiraalvormige secties bevinden. De tertiaire structuur van MB heeft de vorm van een compacte bol, in de verdieping waarvan tussen de F- en E α-helices een actief centrum is.
Ppt% 5C34928-slozhnye_belki_ch1_41.jpg "alt =" (! LANG:> Myoglobinestructuur">!}
Ppt% 5C34928-slozhnye_belki_ch1_42.jpg "alt =" (! TAAL:> Kenmerken van de structuur en werking van het MB actieve centrum. Het MB actieve centrum wordt voornamelijk gevormd door hydrofobe radicalen"> Особенности строения и функционирования активного центра Мв. Активный центр Мв сформирован преимущественно гидрофобными радикалами аминокислот, далеко отстоящими друг от друга в первичной структуре (например, Три39 и Фен138). К активному центру присоединяется плохо растворимые в воде лиганды - гем и О2. Гем - специфический лиганд апоМв.!}
Ppt% 5C34928-slozhnye_belki_ch1_43.jpg "alt =" (! LANG:> Het heem is gebaseerd op vier pyrroolringen verbonden door methhenylbruggen; het Fe2 + -atoom bevindt zich in het midden,"> Основу гема составляют четыре пиррольных кольца, соединенных метенильными мостиками; в центре расположен атом Fe2+, соединенный с атомами азота пиррольных колец четырьмя координационными связями. В активном центре Мв кроме гидрофобных радикалов аминокислот имеются также остатки двух аминокислот с гидрофильными радикалами - Гис Е7 (Гис64) и Гис F8 (Гис93).!}
Ppt% 5C34928-slozhnye_belki_ch1_44.jpg "alt =" (! TAAL:> Zijn F8 vormt een coördinatiebinding met Fe2 + en fixeert de heem stevig in het actieve centrum."> Гис F8 образует координационную связь с Fe2+ и прочно фиксирует гем в активном центре. Гис Е7 необходим для правильной ориентации в активном центре другого лиганда - O2 при его взаимодействии с Fe+2 гема. Микроокружение гема создает условия для прочного, но обратимого связывания O2 с Fe+2 и препятствует попаданию в гидрофобный активный центр воды, что может привести к его окислению в Fе3+.!}
Ppt% 5C34928-slozhnye_belki_ch1_45.jpg "alt =" (! TAAL:> Oligomere structuur van HB en regulatie van de affiniteit van HB voor O2-liganden. Menselijke hemoglobines -"> Олигомерное строение Нв и регуляция сродства Нв к О2 лигандами. Гемоглобины человека - семейство белков, так же как и миоглобин относящиеся к сложным белкам (гемопротеинам). Они имеют тетрамерное строение и содержат две α-цепи, но различаются по строению двух других полипептидных цепей (2α-, 2х-цепи). Строение второй полипептидной цепи определяет особенности функционирования этих форм Нв. Около 98% гемоглобина эритроцитов взрослого человека составляет гемоглобин А (2α-, 2β-цепи). В период внутриутробного развития функционируют два основных типа гемоглобинов: эмбриональный Нв (2α, 2ε), который обнаруживается на ранних этапах развития плода, и гемоглобин F (фетальный) - (2α, 2γ), который приходит на смену раннему гемоглобину плода на шестом месяце внутриутробного развития и только после рождения замещается на Нв А.!}
Ppt% 5C34928-slozhnye_belki_ch1_46.jpg "alt =" (! TAAL:> HB A is een eiwit gerelateerd aan myoglobine (MB), dat wordt aangetroffen in de erytrocyten van een volwassene."> Нв А - белок, родственный миоглобину (Мв), содержится в эритроцитах взрослого человека. Строение его отдельных протомеров аналогично таковому у миоглобина. Вторичная и третичная структуры миоглобина и протомеров гемоглобина очень сходны, несмотря на то что в первичной структуре их полипептидных цепей идентичны только 24 аминокислотных остатка (вторичная структура протомеров гемоглобина, так же как миоглобин, содержит восемь α-спиралей, обозначаемых латинскими буквами от А до Н, а третичная структура имеет вид компактной глобулы). Но в отличие от миоглобина гемоглобин имеет олигомерное строение, состоит из четырех полипептидных цепей, соединенных нековалентными связями.!}
Ppt% 5C34928-slozhnye_belki_ch1_47.jpg "alt =" (! LANG:> Hemoglobine oligomere structuur">!}
Ppt% 5C34928-slozhnye_belki_ch1_48.jpg "alt =" (! TAAL:> Elke HB-protomeer wordt geassocieerd met een niet-eiwitdeel - heem en aangrenzende protomeren."> Каждый протомер Нв связан с небелковой частью - гемом и соседними протомерами. Соединение белковой части Нв с гемом аналогично таковому у миоглобина: в активном центре белка гидрофобные части гема окружены гидрофобными радикалами аминокислот за исключением Гис F8 и Гис Е7, которые расположены по обе стороны от плоскости гема и играют аналогичную роль в функционировании белка и связывании его с кислородом. Кроме того, Гис Е7 выполняет важную дополнительную роль в функционировании Нв. Свободный гем имеет в 25 000 раз более высокое сродство к СО, чем к О2. СО в небольших количествах образуется в организме и, учитывая его высокое сродство к гему, он мог бы нарушать транспорт необходимого для жизни клеток О2. Однако в составе гемоглобина сродство гема к оксиду углерода превышает сродство к О2 всего в 200 раз благодаря наличию в активном центре Гис Е7. Остаток этой аминокислоты создает !} optimale omstandigheden voor binding van heem aan O2 en verzwakt de interactie van heem met CO.
Ppt% 5C34928-slozhnye_belki_ch1_49.jpg "alt ="> ">
Ppt% 5C34928-slozhnye_belki_ch1_50.jpg "alt =" (! LANG:> De heempyrroolringen bevinden zich in hetzelfde vlak en het Fe2 + -ion bevindt zich in de zuurstofvrije toestand Hb"> Пиррольные кольца гема расположены в одной плоскости, а ион Fe2+ в неоксигенированом состоянии Hb выступает над плоскостью на 0,6 А. При присоединении кислорода ион железа погружается в плоскость колец гема. В результате сдвигается и участок полипептидной цепи, нарушаются слабые связи в молекуле Hb и изменяется конформация всей глобулы. Таким образом, присоединение кислорода вызывает изменение пространственной структуры молекулы миоглобина или протомеров гемоглобина.!}
Ppt% 5C34928-slozhnye_belki_ch1_51.jpg "alt =" (! LANG:> Hemoglobine kan zowel in vrije (deoxyhemoglobine) als in zuurstofrijke vorm voorkomen, hechtend aan"> Гемоглобин может существовать как в свободной (дезоксигемоглобин), так и в оксигенированной форме, присоединяя до 4 молекул кислорода. Взаимодействие с кислородом 1-го протомера вызывает изменение его конформации, а также кооперативные конформационные изменения остальных протомеров. Сродство к кислороду возрастает, и присоединение О2 к активному центру 2-го протомера происходит легче, вызывая дальнейшую конформационную перестройку всей молекулы. В результате еще сильнее изменяется структура оставшихся протомеров и их активных центров, взаимодействие с О2 еще больше облегчается. В итоге 4-я молекула кислорода присоединяется к Hb примерно в 300 раз легче, чем 1-я. Так происходит в легких при высоком парциальном давлении кислорода.!}
Ppt% 5C34928-slozhnye_belki_ch1_52.jpg "alt =" (! LANG:> Coöperatieve veranderingen in de conformatie van het hemoglobinemolecuul bij interactie met zuurstof">!}
Ppt% 5C34928-slozhnye_belki_ch1_53.jpg "alt =" (! TAAL:> In weefsels waar het zuurstofgehalte lager is, daarentegen, vergemakkelijkt de splitsing van elk O2-molecuul de afgifte van volgende moleculen."> В тканях, где содержание кислорода ниже, наоборот, отщепление каждой молекулы О2 облегчает освобождение последующих. Таким образом, взаимодействие олигомерного белка гемоглобина с лигандом (О2) в одном центре связывания приводит к изменению конформации всей молекулы и других, пространственно удаленных центров, расположенных на других субъединицах (принцип «домино»). Подобные взаимосвязанные изменения структуры белка называют кооперативными конформационными изменениями. Они характерны для всех олигомерных белков и используются для регуляции их активности.!}
Ppt% 5C34928-slozhnye_belki_ch1_54.jpg "alt =" (! LANG:> De interactie van beide eiwitten (Mb en Hb) met zuurstof hangt af van de partiële druk in"> Взаимодействие обоих белков (Mb и Hb) с кислородом зависит от его парциального давления в тканях. Эта зависимость имеет разный характер, что связано с их особенностями структуры и функционирования. Гемоглобин имеет S-образную кривую насыщения, которая показывает, что субъединицы белка работают кооперативно, и чем больше кислорода они отдают, тем легче идет освобождение остальных молекул О2. Этот процесс зависит от изменения парциального давления кислорода в тканях. График насыщения миоглобина кислородом имеет характер простой гиперболы, т.е. насыщение Mb кислородом происходит быстро и отражает его функцию - обратимое связывание с кислородом, высвобождаемым гемоглобином, и освобождение в случае интенсивной физической нагрузки.!}
Ppt% 5C34928-slozhnye_belki_ch1_55.jpg "alt =" (! LANG:> Myoglobine en hemoglobine zuurstofverzadigingscurves">!}
Ppt% 5C34928-slozhnye_belki_ch1_56.jpg "alt =" (! LANG:> CO2 en H +, gevormd tijdens het katabolisme van organische stoffen, verminderen de affiniteit van hemoglobine voor O2 proportioneel"> CO2 и Н+, образующиеся при катаболизме органических веществ, уменьшают сродство гемоглобина к О2 пропорционально их концентрации. Энергия, необходимая для работы клеток, вырабатывается преимущественно в митохондриях при окислении органических веществ с использованием O2, доставляемого из легких гемоглобином. В результате окисления органических веществ образуются конечные продукты их распада: СО2 и Н2O, количество которых пропорционально интенсивности протекающих процессов окисления. СO2 диффузией попадает из клеток в кровь и проникает в эритроциты, где под действием фермента карбоангидразы превращается в угольную кислоту. Эта слабая кислота диссоциирует на протон и бикарбонат ион. СО2 + Н2О → Н2СО3 → Н+ + НСО3-!}
Ppt% 5C34928-slozhnye_belki_ch1_57.jpg "alt =" (! LANG:> H + -ionen kunnen zich hechten aan His146-radicalen in de β-ketens van hemoglobine, dat wil zeggen in afgelegen gebieden"> Ионы Н+ способны присоединятся к радикалам Гис146 в β-цепях гемоглобина, т.е. в участках, удаленных от гема. Протонирование гемоглобина снижает его сродство к О2, способствует отщеплению О2 от оксиНв, образованию дезоксиНв и увеличивает поступление кислорода в ткани пропорционально количеству образовавшихся протонов. Увеличение количества освобожденного кислорода в зависимости от увеличения концентрации Н+ в эритроцитах называется эффектом Бора (по имени датского физиолога Христиана Бора, впервые открывшего этот эффект). В легких высокое парциальное давление кислорода способствует его связыванию с дезоксиНв, что уменьшает сродство белка к Н+. Освободившиеся протоны под действием карбоангидразы взаимодействуют с бикарбонатами с образованием СО2 и Н2О!}
Ppt% 5C34928-slozhnye_belki_ch1_58.jpg "alt =" (! LANG:> Afhankelijkheid van de Hb-affiniteit voor O2 van de concentratie van CO2 en protonen (Bohr-effect): A -"> Зависимость сродства Нв к О2 от концентрации СО2 и протонов (эффект Бора): А - влияние концентрации СО2 и Н+ на высвобождение О2 из комплекса с Нв (эффект Бора); Б - оксигенирование дезоксигемоглобина в легких, образование и выделение СО2.!}
Ppt% 5C34928-slozhnye_belki_ch1_59.jpg "alt =" (! LANG:> Het resulterende CO2 komt de alveolaire ruimte binnen en wordt verwijderd met uitgeademde lucht."> Образовавшийся СО2 поступает в альвеолярное пространство и удаляется с выдыхаемым воздухом. Таким образом, количество высвобождаемого гемоглобином кислорода в тканях регулируется продуктами катаболизма органических веществ: чем интенсивнее распад веществ, например при физических нагрузках, тем выше концентрация СО2 и Н+ и тем больше кислорода получают ткани в результате уменьшения сродства Нв к О2.!}
Ppt% 5C34928-slozhnye_belki_ch1_60.jpg "alt =" (! LANG:> De verandering in de functionele activiteit van het eiwit bij interactie met andere liganden als gevolg van conformationele veranderingen wordt allosterisch genoemd"> Изменение функциональной активности белка при взаимодействии с другими лигандами вследствие конформационных изменений называется аллостерической регуляцией, а соединения-регуляторы - аллостерическими лигандами или эффекторами. Способность к аллостерической регуляции характерна, как правило, для олигомерных белков, т.е. для проявления аллостерического эффекта необходимо взаимодействие протомеров. При воздействии аллостерических лигандов белки меняют свою конформацию (в том числе и активного центра) и функцию.!}
Ppt% 5C34928-slozhnye_belki_ch1_61.jpg "alt =" (! LANG:> Allosterische regulatie van HB's affiniteit voor O2 door het ligand 2,3-bis-fosfoglyceraat. In erytrocyten van het product"> Аллостерическая регуляция сродства Нв к О2 лигандом - 2,3-бис-фосфоглицератом. В эритроцитах из продукта окисления глюкозы - 1,3-бисфосфоглицерата синтезируется аллостерический лиганд гемоглобина - 2,3-бисфосфоглицерат (2,3-БФГ). В нормальных условиях концентрация 2,3-БФГ высокая и сравнима с концентрацией Нв. 2,3-БФГ имеет сильный отрицательный заряд (-5).!}
Ppt% 5C34928-slozhnye_belki_ch1_62.jpg "alt =" (! TAAL:> In het midden van het tetramere hemoglobinemolecuul bevindt zich een holte gevormd door de aminozuurresiduen van alle vier de protomeren."> В центре тетрамерной молекулы гемоглобина находится полость. Ее образуют аминокислотные остатки всех четырех протомеров. В капиллярах тканей протонирование Нв (эффект Бора) приводит к разрыву связи между железом гема и О2. В молекуле дезоксигемоглобина по сравнению с оксигемоглобином возникают дополнительные ионные связи, соединяющие протомеры, вследствие чего размеры центральной полости по сравнению с оксигемоглобином увеличиваются. Центральная полость является местом присоединения 2,3-БФГ к гемоглобину. БФГ поступает в полость дезоксигемоглобина. 2,3-БФГ взаимодействует с гемоглобином в участке, удаленном от активных центров белка и относится к аллостерическим (регуляторным) лигандам, а центральная полость Нв является аллостерическим центром. 2,3-БФГ имеет сильный отрицательный заряд и взаимодействует с положительно заряженными группами двух β-цепей Нв. При этом его сродство к О2 снижается в 26 раз. В результате происходит высвобождение кислорода в капиллярах ткани при низком парциальном давлении О2. В легких высокое парциальное давление О2, наоборот, приводит к оксигенированию Нв и освобождению БФГ.!}
Ppt% 5C34928-slozhnye_belki_ch1_63.jpg "alt =" (! LANG:> Het BFG-bindingscentrum bevindt zich in een positief geladen holte tussen de 4 hemoglobine-protomeren."> Центр связывания БФГ находится в положительно заряженной полости между 4 протомерами гемоглобина. Взаимодействие БФГ с центром связывания изменяет конформацию α- и β-протомеров НЬ и их активных центров. Сродство НЬ к молекулам О2 снижается и кислород высвобождается в ткани. В легких при высоком парциальном давлении О2 активные центры гемоглобина насыщаются за счет изменения конформации и БФГ вытесняется из аллостерического центра!}
Ppt% 5C34928-slozhnye_belki_ch1_64.jpg "alt ="> ">
Ppt% 5C34928-slozhnye_belki_ch1_65.jpg "alt =" (! TAAL:> Oligomere eiwitten hebben dus nieuwe eigenschappen in vergelijking met monomere eiwitten."> Таким образом, олигомерные белки обладают новыми по сравнению с мономерными белками свойствами. Присоединение лигандов на участках, пространственно удаленных друг от друга (аллостерических), способно вызывать конформационные изменения во всей белковой молекуле. Благодаря взаимодействию с регуляторными лигандами происходит изменение конформации и адаптация функции белковой молекулы к изменениям окружающей среды.!}
Ppt% 5C34928-slozhnye_belki_ch1_66.jpg "alt =" (! TAAL:> Ongeveer 15% van de koolstofdioxide die in het bloed aanwezig is, wordt gedragen door hemoglobinemoleculen."> Около 15% углекислого газа, присутствующего в крови, переносится молекулами гемоглобина. В тканях часть молекул углекислого газа может присоединится к каждому протомеру молекулы гемоглобина, при этом снижается сродство Hb к кислороду. В легких, наоборот, из-за высокого парциального давления кислорода, О2 связывается с Hb, а СО2 высвобождается.!}
Ppt% 5C34928-slozhnye_belki_ch1_67.jpg "alt ="> ">
Ppt% 5C34928-slozhnye_belki_ch1_68.jpg "alt =" (! LANG:> In het hemoglobine S-molecuul (het zogenaamde abnormale hemoglobine) werden 2 β-ketens gemuteerd, waarin"> В молекуле гемоглобина S (так назван аномальный гемоглобин) мутантными оказались 2 β-цепи, в которых глутамат, высокополярная отрицательно заряженная аминокислота в положении 6 была заменена валином, содержащим гидрофобный радикал.!}
Modulestructuur | Thema's |
Modulaire eenheid 1 | 1.1. Structurele organisatie van eiwitten. Stadia van de vorming van natieve eiwitconformatie 1.2. Grondbeginselen van de werking van eiwitten. Geneesmiddelen als liganden die de eiwitfunctie beïnvloeden 1.3. Denaturatie van eiwitten en de mogelijkheid van hun spontane vernieuwing |
Modulaire eenheid 2 | 1.4. Kenmerken van de structuur en werking van oligomere eiwitten naar het voorbeeld van hemoglobine 1.5. Handhaving van de natieve conformatie van eiwitten onder celomstandigheden 1.6. Verscheidenheid aan eiwitten. Eiwitfamilies zoals geïllustreerd door immunoglobulinen 1.7. Fysisch-chemische eigenschappen van eiwitten en methoden voor hun scheiding |
Modulaire eenheid 1 STRUCTURELE ORGANISATIE VAN MONOMERISCHE EIWITTEN EN DE BASIS VAN HUN WERKING
Leerdoelen In staat zijn om:
1. Gebruik kennis over de kenmerken van de structuur van eiwitten en de afhankelijkheid van de functies van eiwitten van hun structuur om de mechanismen van ontwikkeling van erfelijke en verworven proteïnopathieën te begrijpen.
2. Verklaar mechanismen therapeutische actie sommige medicijnen als liganden die interageren met eiwitten en hun activiteit veranderen.
3. Gebruik kennis over de structuur en conformationele labiliteit van eiwitten om hun structurele en functionele instabiliteit en neiging tot denaturatie onder veranderende omstandigheden te begrijpen.
4. Leg het gebruik uit van denaturerende middelen als middelen voor het steriliseren van medisch materiaal en instrumenten, evenals antiseptica.
Weten:
1. Niveaus van de structurele organisatie van eiwitten.
2. De waarde van de primaire structuur van eiwitten, die hun structurele en functionele diversiteit bepaalt.
3. Het mechanisme van vorming van een actief centrum in eiwitten en de specifieke interactie met de ligand, die ten grondslag ligt aan de werking van eiwitten.
4. Voorbeelden van de invloed van exogene liganden (medicijnen, toxines, vergiften) op de conformatie en functionele activiteit van eiwitten.
5. Oorzaken en gevolgen van eiwitdenaturatie, factoren die denaturatie veroorzaken.
6. Voorbeelden van het gebruik van denaturerende factoren in de geneeskunde als antiseptica en middelen voor het steriliseren van medische instrumenten.
ONDERWERP 1.1. STRUCTURELE ORGANISATIE VAN EIWITTEN. STAPPEN VAN VORMING VAN INHEEMSE
EIWIT CONFORMATIES
Eiwitten zijn polymere moleculen met slechts 20 -aminozuren als monomeren. De set en volgorde waarin aminozuren in een eiwit worden gecombineerd, wordt bepaald door de structuur van genen in het DNA van individuen. Elk eiwit vervult, in overeenstemming met zijn specifieke structuur, zijn eigen functie. De set eiwitten van een bepaald organisme bepaalt de fenotypische kenmerken ervan, evenals de aanwezigheid van erfelijke ziekten of een aanleg voor hun ontwikkeling.
1. Aminozuren waaruit eiwitten bestaan. Peptide binding. Eiwitten zijn polymeren die zijn opgebouwd uit monomeren - 20 -aminozuren, waarvan de algemene formule is:
Aminozuren verschillen in structuur, grootte en fysisch-chemische eigenschappen van radicalen die aan het α-koolstofatoom zijn gehecht. De functionele groepen van aminozuren bepalen de eigenschappen van de eigenschappen van verschillende α-aminozuren. De radicalen die in -aminozuren worden gevonden, kunnen worden onderverdeeld in verschillende groepen:
Proline, in tegenstelling tot andere 19 eiwitmonomeren, is het geen aminozuur, maar een iminozuur, het radicaal in proline is gekoppeld aan zowel het α-koolstofatoom als de iminogroep
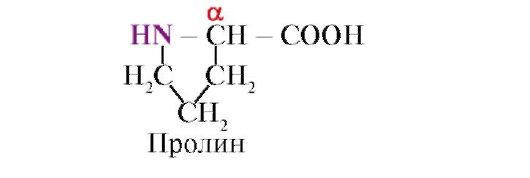 Aminozuren verschillen in hun oplosbaarheid in water. Dit komt door het vermogen van radicalen om te interageren met water (hydraat).
Aminozuren verschillen in hun oplosbaarheid in water. Dit komt door het vermogen van radicalen om te interageren met water (hydraat).
NAAR hydrofiel omvat radicalen die anionische, kationische en polaire ongeladen functionele groepen bevatten.
NAAR hydrofoob omvat radicalen die methylgroepen, alifatische ketens of cycli bevatten.
2. Peptidebindingen verbinden aminozuren tot peptiden. Bij de synthese van een peptide interageert de α-carboxylgroep van het ene aminozuur met de -aminogroep van een ander aminozuur om peptide binding:
Eiwitten zijn polypeptiden, d.w.z. lineaire polymeren van -aminozuren verbonden door een peptidebinding (Fig.1.1.)
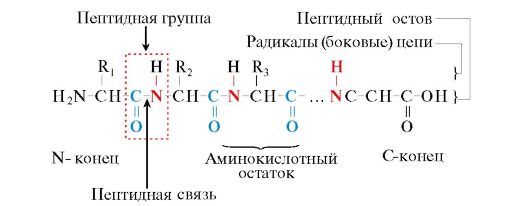 Rijst. 1.1. Termen die worden gebruikt om de structuur van peptiden te beschrijven
Rijst. 1.1. Termen die worden gebruikt om de structuur van peptiden te beschrijven
Monomeren van aminozuren waaruit polypeptiden bestaan, worden aminozuurresten. Een keten van herhalende groepen - NH-CH-CO- formulieren peptide ruggengraat. Een aminozuurrest met een vrije -aminogroep wordt N-terminaal genoemd en een aminozuurrest met een vrije α-carboxylgroep wordt C-terminaal genoemd. De peptiden worden geschreven en gelezen van het N-uiteinde naar het C-uiteinde.
De peptidebinding gevormd door de iminogroep van proline verschilt van andere peptidebindingen: het stikstofatoom van de peptidegroep mist waterstof,
in plaats daarvan is er een binding met de radicaal, waardoor één kant van de cyclus wordt opgenomen in de peptide-ruggengraat:
 Peptiden verschillen in aminozuursamenstelling, het aantal aminozuren en de volgorde waarin aminozuren worden gecombineerd, Ser-Ala-Glu-Gis en Gis-Glu-Ala-Ser zijn bijvoorbeeld twee verschillende peptiden.
Peptiden verschillen in aminozuursamenstelling, het aantal aminozuren en de volgorde waarin aminozuren worden gecombineerd, Ser-Ala-Glu-Gis en Gis-Glu-Ala-Ser zijn bijvoorbeeld twee verschillende peptiden.
Peptidebindingen zijn erg sterk en hun chemische niet-enzymatische hydrolyse vereist strikte voorwaarden: het geanalyseerde eiwit wordt gehydrolyseerd in geconcentreerd zoutzuur bij een temperatuur van ongeveer 110 ° gedurende 24 uur. In een levende cel kunnen peptidebindingen worden verbroken met behulp van Proteolytische enzymen, genaamd proteasen of peptide hydrolasen.
3. Primaire structuur van eiwitten. Aminozuurresten in de peptideketens van verschillende eiwitten wisselen elkaar niet willekeurig af, maar zijn in een bepaalde volgorde gerangschikt. De lineaire volgorde of de volgorde van afwisseling van aminozuurresten in de polypeptideketen wordt genoemd de primaire structuur van het eiwit.
De primaire structuur van elk afzonderlijk eiwit wordt gecodeerd in een DNA-molecuul (in een gebied dat een genoom wordt genoemd) en wordt gerealiseerd tijdens transcriptie (herschrijven van informatie over mRNA) en translatie (synthese van de primaire structuur van een eiwit). Bijgevolg is de primaire structuur van eiwitten van een individuele persoon informatie die erfelijk wordt overgedragen van ouders op kinderen, die de structurele kenmerken van de eiwitten van een bepaald organisme bepaalt, waarvan de functie van bestaande eiwitten afhangt (Fig. 1.2.).
 Rijst. 1.2. De relatie tussen het genotype en de conformatie van eiwitten gesynthetiseerd in het lichaam van een individu
Rijst. 1.2. De relatie tussen het genotype en de conformatie van eiwitten gesynthetiseerd in het lichaam van een individu
Elk van de ongeveer 100.000 individuele eiwitten in het menselijk lichaam heeft uniek primaire structuur. De moleculen van één type eiwit (bijvoorbeeld albumine) hebben dezelfde afwisseling van aminozuurresiduen, wat albumine onderscheidt van elk ander afzonderlijk eiwit.
De volgorde van aminozuurresten in de peptideketen kan worden beschouwd als een vorm van informatieregistratie. Deze informatie bepaalt de ruimtelijke vouwing van de lineaire peptideketen in een compactere driedimensionale structuur genaamd conformatie eekhoorn. Het proces van het vormen van een functioneel actieve eiwitconformatie heet vouwen.
4. Conformatie van eiwitten. Vrije rotatie in de peptideruggengraat is mogelijk tussen het stikstofatoom van de peptidegroep en het aangrenzende -koolstofatoom, evenals tussen het α-koolstofatoom en het koolstofatoom van de carbonylgroep. Door de interactie van functionele groepen van aminozuurresiduen kan de primaire structuur van eiwitten complexere ruimtelijke structuren krijgen. In bolvormige eiwitten zijn er twee hoofdniveaus van vouwing van de conformatie van peptideketens: ondergeschikt en tertiaire structuur.
Secundaire structuur van eiwitten is een ruimtelijke structuur gevormd als gevolg van de vorming van waterstofbruggen tussen de functionele groepen -C = O en -NH- van de peptideruggengraat. In dit geval kan de peptideketen regelmatige structuren van twee typen krijgen: α-helix en β-structuren.
V α-helix waterstofbruggen worden gevormd tussen het zuurstofatoom van de carbonylgroep en de waterstof van de amidestikstof van het 4e aminozuur ervan; zijketens van aminozuurresiduen
bevinden zich langs de omtrek van de spiraal, zonder deel te nemen aan de vorming van de secundaire structuur (Fig. 1.3.).
Omvangrijke radicalen of radicalen met dezelfde lading voorkomen de vorming van de α-helix. Het prolineresidu, dat een ringstructuur heeft, onderbreekt de α-helix, omdat het door de afwezigheid van waterstof aan het stikstofatoom in de peptideketen onmogelijk is om een waterstofbinding te vormen. De binding tussen stikstof en het α-koolstofatoom maakt deel uit van de prolinecyclus; daarom krijgt de peptideruggengraat op dit punt een bocht.
β-structuur vormt zich tussen de lineaire gebieden van de peptideruggengraat van één polypeptideketen, waardoor gevouwen structuren worden gevormd. Polypeptideketens of delen daarvan kunnen zich vormen parallel of antiparallelle -structuren. In het eerste geval vallen de N- en C-uiteinden van de interagerende peptideketens samen, en in het tweede geval hebben ze de tegenovergestelde richting (Fig. 1.4).
Rijst. 1.3. Secundaire structuur van het eiwit - α-helix
 Rijst. 1.4. Parallelle en antiparallelle β-gevouwen structuren
Rijst. 1.4. Parallelle en antiparallelle β-gevouwen structuren
β-structuren zijn aangegeven met brede pijlen: A - Antiparallelle β-structuur. B - Parallelle β-gevouwen structuren
In sommige eiwitten kunnen β-structuren worden gevormd door de vorming van waterstofbruggen tussen de atomen van de peptideruggengraat van verschillende polypeptideketens.
Ook gevonden in eiwitten gebieden met onregelmatige secundaire structuur, waaronder bochten, lussen, bochten van de ruggengraat van het polypeptide. Ze bevinden zich vaak op plaatsen waar de richting van de peptideketen verandert, bijvoorbeeld tijdens de vorming van een parallelle β-sheetstructuur.
Afhankelijk van de aanwezigheid van α-helices en β-structuren, kunnen bolvormige eiwitten worden onderverdeeld in vier categorieën.
Rijst. 1.5. Secundaire structuur van myoglobine (A) en β-ketens van hemoglobine (B), met acht α-helices

 Rijst. 1.6. Secundaire structuur van triosefosfaatisomerase en pyruvaatkinasedomein
Rijst. 1.6. Secundaire structuur van triosefosfaatisomerase en pyruvaatkinasedomein
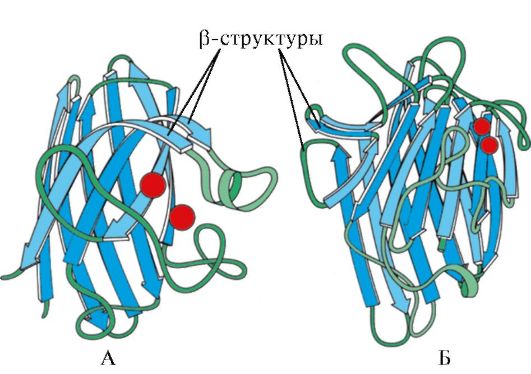 Rijst. 1.7. Secundaire structuur van het constante domein van immunoglobuline (A) en het enzym superoxide dismutase (B)
Rijst. 1.7. Secundaire structuur van het constante domein van immunoglobuline (A) en het enzym superoxide dismutase (B)
V vierde categorie inclusief eiwitten die een kleine hoeveelheid reguliere secundaire structuren bevatten. Deze omvatten kleine, cysteïnerijke eiwitten of metalloproteïnen.
Tertiaire eiwitstructuur- het type conformatie dat wordt gevormd door interacties tussen aminozuurradicalen, die zich op aanzienlijke afstand van elkaar in de peptideketen kunnen bevinden. In dit geval vormen de meeste eiwitten een ruimtelijke structuur die lijkt op een bolletje (bolvormige eiwitten).
Omdat hydrofobe aminozuurradicalen de neiging hebben om te combineren met behulp van de zogenaamde hydrofobe interacties en intermoleculaire van der Waals-krachten, wordt een dichte hydrofobe kern gevormd in het eiwitbolletje. Hydrofiele geïoniseerde en niet-geïoniseerde radicalen bevinden zich voornamelijk op het oppervlak van het eiwit en bepalen de oplosbaarheid in water.
 Rijst. 1.8. Soorten bindingen die ontstaan tussen aminozuurradicalen tijdens de vorming van de tertiaire structuur van een eiwit
Rijst. 1.8. Soorten bindingen die ontstaan tussen aminozuurradicalen tijdens de vorming van de tertiaire structuur van een eiwit
1 - ionbinding- komt voor tussen positief en negatief geladen functionele groepen;
2 - waterstofbinding- komt voor tussen een ongeladen hydrofiele groep en een andere hydrofiele groep;
3 - hydrofobe interacties- ontstaan tussen hydrofobe radicalen;
4 - disulfidebinding- gevormd door de oxidatie van SH-groepen van cysteïneresiduen en hun interactie met elkaar
Hydrofiele aminozuurresiduen die in de hydrofobe kern zijn opgesloten, kunnen met elkaar interageren met behulp van ionisch en waterstofbruggen(afb. 1.8).
Ionische en waterstofbruggen, evenals hydrofobe interacties, behoren tot de zwakke: hun energie is iets hoger dan de energie van de thermische beweging van moleculen op kamertemperatuur... Eiwitconformatie wordt gehandhaafd door de opkomst van veel van deze zwakke bindingen. Omdat de atomen waaruit het eiwit bestaat constant in beweging zijn, kunnen sommige zwakke bindingen worden verbroken en andere worden gevormd, wat leidt tot kleine verplaatsingen van afzonderlijke delen van de polypeptideketen. Deze eigenschap van eiwitten om van conformatie te veranderen als gevolg van het verbreken van sommige en de vorming van andere zwakke bindingen wordt genoemd conformationele labiliteit.
Het menselijk lichaam heeft systemen die ondersteuning bieden homeostase- de constantheid van het interne milieu binnen bepaalde voor een gezond organisme toelaatbare grenzen. Onder homeostase-omstandigheden verstoren kleine veranderingen in conformatie de algehele structuur en functie van eiwitten niet. De functioneel actieve conformatie van een eiwit heet inheemse conformatie. Een verandering in de interne omgeving (bijvoorbeeld de concentratie van glucose, Ca-ionen, protonen, enz.) leidt tot een verandering in conformatie en disfunctie van eiwitten.
De tertiaire structuur van sommige eiwitten is gestabiliseerd Disulfide verbindingen, gevormd door de interactie van -SH-groepen van twee residuen
 Rijst. 1.9. Vorming van een disulfidebinding in een eiwitmolecuul
Rijst. 1.9. Vorming van een disulfidebinding in een eiwitmolecuul
cysteïne (Fig. 1.9). De meeste intracellulaire eiwitten hebben geen covalente disulfidebindingen in de tertiaire structuur. Hun aanwezigheid is kenmerkend voor eiwitten die door de cel worden uitgescheiden, wat zorgt voor een grotere stabiliteit in extracellulaire omstandigheden. Dus disulfidebindingen worden gevonden in insuline- en immunoglobulinemoleculen.
Insuline- een eiwithormoon dat in de β-cellen van de pancreas wordt gesynthetiseerd en in het bloed wordt uitgescheiden als reactie op een verhoging van de glucoseconcentratie in het bloed. In de insulinestructuur zijn er twee disulfidebindingen die de polypeptide A- en B-ketens verbinden, en één disulfidebinding binnen de A-keten (Fig. 1.10).
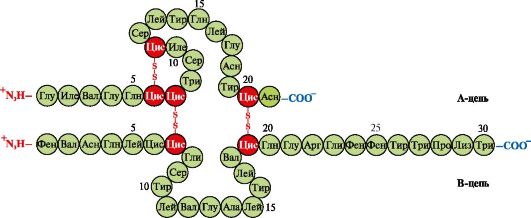 Rijst. 1.10. Disulfidebindingen in de structuur van insuline
Rijst. 1.10. Disulfidebindingen in de structuur van insuline
5. Supersecundaire structuur van eiwitten. In eiwitten die verschillen in hun primaire structuur en functies, soortgelijke combinaties en rangschikking van secundaire structuren, die supersecundaire structuur worden genoemd. Het neemt een tussenpositie in tussen de secundaire en tertiaire structuren, aangezien dit een specifieke combinatie is van elementen van de secundaire structuur bij de vorming van de tertiaire structuur van een eiwit. Supersecundaire structuren hebben specifieke namen zoals "α-helix-turn-a-helix", "leucine zipper", "zinkvingers", enz. Dergelijke supersecundaire structuren zijn kenmerkend voor DNA-bindende eiwitten.
Leucine rits. Dit soort supersecundaire structuur wordt gebruikt om twee eiwitten met elkaar te verbinden. Op het oppervlak van interagerende eiwitten bevinden zich a-helixgebieden die ten minste vier leucineresiduen bevatten. Leucineresiduen in de α-helix bevinden zich zes aminozuren van elkaar. Aangezien elke winding van de α-helix 3,6 aminozuurresiduen bevat, bevinden leucineradicalen zich op het oppervlak van elke tweede winding. Leucineresiduen van de α-helix van één eiwit kunnen interageren met leucineresiduen van een ander eiwit (hydrofobe interacties), waardoor ze met elkaar worden verbonden (Fig. 1.11.). Veel DNA-bindende eiwitten functioneren als onderdeel van oligomere complexen, waarbij individuele subeenheden aan elkaar binden met "leucinebevestigingsmiddelen".
 Rijst. 1.11. "Leucine-ritssluiting" tussen de α-helixgebieden van twee eiwitten
Rijst. 1.11. "Leucine-ritssluiting" tussen de α-helixgebieden van twee eiwitten
Een voorbeeld van dergelijke eiwitten zijn histonen. histonen- nucleaire eiwitten, die een groot aantal positief geladen aminozuren bevatten - arginine en lysine (tot 80%). Histonmoleculen worden gecombineerd tot oligomere complexen die acht monomeren bevatten met behulp van "leucinebevestigingsmiddelen", ondanks de aanzienlijke lading met dezelfde naam van deze moleculen.
"Zink vinger"- een variant van de supersecundaire structuur, kenmerkend voor DNA-bindende eiwitten, heeft de vorm van een langwerpig fragment op het eiwitoppervlak en bevat ongeveer 20 aminozuurresten (Fig. 1.12). De vorm van de "langwerpige vinger" wordt ondersteund door een zinkatoom gekoppeld aan radicalen van vier aminozuren - twee cysteïneresiduen en twee histidineresiduen. In sommige gevallen zijn er in plaats van histidineresten cysteïneresten. Twee dicht bij elkaar gelegen cysteïneresiduen worden van de andere twee Gisili-residuen gescheiden door een sequentie van ongeveer 12 aminozuurresiduen. Dit gebied van het eiwit vormt een a-helix, waarvan de radicalen specifiek kunnen binden aan de regulerende gebieden van de grote groef van DNA. De bindingsspecificiteit van een individu
 Rijst. 1.12. De primaire structuur van het gebied van DNA-bindende eiwitten die de structuur van de "zinkvinger" vormen (de letters geven de aminozuren aan waaruit deze structuur bestaat)
Rijst. 1.12. De primaire structuur van het gebied van DNA-bindende eiwitten die de structuur van de "zinkvinger" vormen (de letters geven de aminozuren aan waaruit deze structuur bestaat)
regulerend DNA-bindend eiwit hangt af van de sequentie van aminozuurresiduen die zich in de "zinkvinger" bevinden. Dergelijke structuren bevatten met name receptoren voor steroïde hormonen die betrokken zijn bij de regulatie van transcriptie (het lezen van informatie van DNA naar RNA).
ONDERWERP 1.2. BASIS VAN WERKING VAN EIWITTEN. GENEESMIDDELEN ALS LIGANDEN DIE DE FUNCTIE VAN EIWITTEN BENVLOEDEN
1. Het actieve centrum van het eiwit en zijn interactie met het ligand. Tijdens de vorming van een tertiaire structuur op het oppervlak van een functioneel actief eiwit, meestal in een depressie, wordt een gebied gevormd dat wordt gevormd door aminozuurradicalen die ver van elkaar verwijderd zijn in de primaire structuur. Deze site, die een unieke structuur heeft voor een bepaald eiwit en in staat is om specifiek een interactie aan te gaan met een bepaald molecuul of een groep vergelijkbare moleculen, wordt de eiwit-ligandbindingsplaats of actieve plaats genoemd. Liganden zijn moleculen die interageren met eiwitten.
Hoge specificiteit de interactie van het eiwit met het ligand wordt geleverd door de complementariteit van de structuur van het actieve centrum met de structuur van het ligand.
complementariteit is de ruimtelijke en chemische correspondentie van op elkaar inwerkende oppervlakken. Het actieve centrum moet niet alleen ruimtelijk overeenkomen met het ligand dat erin is opgenomen, maar er moeten ook bindingen (ionische, waterstof en ook hydrofobe interacties) worden gevormd tussen de functionele groepen van de radicalen die in het actieve centrum zijn opgenomen en het ligand, dat het ligand bevat in het actieve centrum (Fig. 1.13).
 Rijst. 1.13. Complementaire interactie van een eiwit met een ligand
Rijst. 1.13. Complementaire interactie van een eiwit met een ligand
Sommige liganden spelen, door zich te hechten aan het actieve centrum van het eiwit, een ondersteunende rol bij het functioneren van eiwitten. Dergelijke liganden worden cofactoren genoemd en eiwitten die een niet-eiwitgedeelte bevatten, worden genoemd complexe eiwitten(in tegenstelling tot eenvoudige eiwitten, die alleen uit het eiwitgedeelte bestaan). Het niet-eiwitdeel, stevig aan het eiwit vastgehecht, heet prothetische groep. De samenstelling van myoglobine, hemoglobine en cytochromen bevat bijvoorbeeld een prothetische groep die stevig is bevestigd aan het actieve centrum - heem, dat een ijzerion bevat. De complexe eiwitten die heem bevatten, worden hemoproteïnen genoemd.
Wanneer specifieke liganden aan eiwitten worden gehecht, komt de functie van deze eiwitten tot uiting. Zo manifesteert albumine, het belangrijkste eiwit in bloedplasma, zijn transportfunctie door hydrofobe liganden aan het actieve centrum te hechten, zoals vetzuren, bilirubine, sommige medicijnen, enz. (Fig. 1.14)
Liganden die een interactie aangaan met de driedimensionale structuur van de peptideketen kunnen niet alleen organische en anorganische moleculen met een laag molecuulgewicht zijn, maar ook macromoleculen:
DNA (de bovenstaande voorbeelden met DNA-bindende eiwitten);
Polysachariden;
 Rijst. 1.14. Relatie tussen genotype en fenotype
Rijst. 1.14. Relatie tussen genotype en fenotype
De unieke primaire structuur van menselijke eiwitten, gecodeerd in het DNA-molecuul, wordt in cellen gerealiseerd in de vorm van een unieke conformatie, structuur van het actieve centrum en functies van eiwitten
In deze gevallen herkent het eiwit een specifieke ligandplaats die evenredig is met en complementair is aan de bindingsplaats. Op het oppervlak van hepatocyten bevinden zich dus receptoreiwitten voor het hormoon insuline, dat ook een eiwitstructuur heeft. De interactie van insuline met de receptor veroorzaakt een verandering in de conformatie en activering van signaalsystemen, wat leidt tot de ophoping van voedingsstoffen in hepatocyten na het eten.
Op deze manier, de werking van eiwitten is gebaseerd op de specifieke interactie van het actieve centrum van het eiwit met het ligand.
2. Domeinstructuur en zijn rol in het functioneren van eiwitten. Lange polypeptideketens van bolvormige eiwitten vouwen vaak in verschillende compacte, relatief onafhankelijke regio's. Ze hebben een onafhankelijke tertiaire structuur, vergelijkbaar met die van bolvormige eiwitten, en worden domeinen. Door de domeinstructuur van eiwitten wordt hun tertiaire structuur gemakkelijker gevormd.
In domeineiwitten bevinden ligandbindingsplaatsen zich vaak tussen domeinen. Trypsine is dus een proteolytisch enzym dat wordt geproduceerd door het exocriene deel van de pancreas en dat nodig is voor de vertering van voedseleiwitten. Het heeft een structuur met twee domeinen en de bindingsplaats voor trypsine met zijn ligand, een voedingseiwit, bevindt zich in de groef tussen de twee domeinen. In het actieve centrum worden omstandigheden gecreëerd die nodig zijn voor de effectieve binding van een specifieke plaats van het voedingseiwit en de hydrolyse van zijn peptidebindingen.
Wanneer het actieve centrum interageert met het ligand, kunnen verschillende domeinen in een eiwit ten opzichte van elkaar bewegen (Fig. 1.15).
Hexokinase- een enzym dat de fosforylering van glucose katalyseert door middel van ATP. De actieve plaats van het enzym bevindt zich in de spleet tussen de twee domeinen. Wanneer hexokinase aan glucose bindt, worden de omringende domeinen gesloten en wordt het substraat opgesloten, waar fosforylering plaatsvindt (zie Fig. 1.15).
 Rijst. 1.15. Binding van hexokinasedomeinen aan glucose
Rijst. 1.15. Binding van hexokinasedomeinen aan glucose
In sommige eiwitten voeren domeinen onafhankelijke functies uit door te binden aan verschillende liganden. Deze eiwitten worden multifunctioneel genoemd.
3. Geneesmiddelen - liganden die de functie van eiwitten beïnvloeden. De interactie van eiwitten met liganden is specifiek. Vanwege de conformationele labiliteit van het eiwit en zijn actieve centrum, is het echter mogelijk om een andere stof te selecteren die ook een interactie zou kunnen aangaan met het eiwit in het actieve centrum of een ander deel van het molecuul.
Een stof die qua structuur vergelijkbaar is met een natuurlijk ligand heet structurele analoog van het ligand of een onnatuurlijk ligand. Het interageert ook met een eiwit in de actieve plaats. De structurele analoog van het ligand kan beide de functie van het eiwit versterken (agonist), dus verminder het (antagonist). Het ligand en zijn structurele analogen concurreren met elkaar voor binding aan het eiwit in hetzelfde centrum. Dergelijke stoffen worden genoemd concurrerende modulatoren(regulatoren) van eiwitfuncties. Veel medicijnen werken als eiwitremmers. Sommigen van hen worden verkregen door chemische modificatie van natuurlijke liganden. Eiwitremmers kunnen medicijnen en vergiften zijn.
Atropine is een competitieve remmer van M-cholinerge receptoren. Acetylcholine is een neurotransmitter voor de overdracht van zenuwimpulsen via cholinerge synapsen. Om excitatie uit te voeren, moet acetylcholine dat vrijkomt in de synaptische spleet een interactie aangaan met een eiwit - de receptor van het postsynaptische membraan. Twee soorten gevonden cholinerge receptoren:
M-receptor, naast acetylcholine, interageert het selectief met muscarine (vliegenzwamtoxine). M - cholinerge receptoren zijn aanwezig op gladde spieren en veroorzaken bij interactie met acetylcholine hun samentrekking;
H-receptor, specifiek binden aan nicotine. H-cholinerge receptoren worden gevonden in de synapsen van dwarsgestreepte skeletspieren.
specifieke remmer M-cholinerge receptoren is atropine. Het wordt gevonden in belladonna- en bilkruidplanten.
 Atropine heeft in de structuur functionele groepen die vergelijkbaar zijn met acetylcholine en hun ruimtelijke rangschikking, daarom behoort het tot competitieve remmers van M-cholinerge receptoren. Aangezien de binding van acetylcholine aan M-cholinerge receptoren contractie van gladde spieren veroorzaakt, wordt atropine gebruikt als een geneesmiddel dat spierspasmen verlicht. (krampstillend). Het is dus bekend dat het gebruik van atropine de oogspieren ontspant bij het bekijken van de fundus, en ook om spasmen bij gastro-intestinale koliek te verlichten. M-cholinerge receptoren zijn ook aanwezig in de centrale zenuwstelsel(centraal zenuwstelsel), daarom kunnen hoge doses atropine een ongewenste reactie van het centrale zenuwstelsel veroorzaken: motorische en mentale agitatie, hallucinaties, convulsies.
Atropine heeft in de structuur functionele groepen die vergelijkbaar zijn met acetylcholine en hun ruimtelijke rangschikking, daarom behoort het tot competitieve remmers van M-cholinerge receptoren. Aangezien de binding van acetylcholine aan M-cholinerge receptoren contractie van gladde spieren veroorzaakt, wordt atropine gebruikt als een geneesmiddel dat spierspasmen verlicht. (krampstillend). Het is dus bekend dat het gebruik van atropine de oogspieren ontspant bij het bekijken van de fundus, en ook om spasmen bij gastro-intestinale koliek te verlichten. M-cholinerge receptoren zijn ook aanwezig in de centrale zenuwstelsel(centraal zenuwstelsel), daarom kunnen hoge doses atropine een ongewenste reactie van het centrale zenuwstelsel veroorzaken: motorische en mentale agitatie, hallucinaties, convulsies.
Ditilin is een competitieve agonist van H-cholinerge receptoren die de functie van neuromusculaire synapsen remt.
 Neuromusculaire synapsen van skeletspieren bevatten H-cholinerge receptoren. Hun interactie met acetylcholine leidt tot spiercontracties. Sommige chirurgische procedures, evenals endoscopische onderzoeken, gebruiken medicijnen die ontspanning van skeletspieren veroorzaken (spierverslappers). Deze omvatten ditiline, een structureel analoog van acetylcholine. Het hecht zich aan H-cholinerge receptoren, maar in tegenstelling tot acetylcholine wordt het zeer langzaam vernietigd door het enzym acetylcholinesterase. Als gevolg van langdurige opening van ionenkanalen en aanhoudende depolarisatie van het membraan, wordt de geleiding van zenuwimpulsen verstoord en treedt spierontspanning op. Aanvankelijk werden deze eigenschappen gevonden in curare-gif, daarom worden dergelijke medicijnen genoemd curariform.
Neuromusculaire synapsen van skeletspieren bevatten H-cholinerge receptoren. Hun interactie met acetylcholine leidt tot spiercontracties. Sommige chirurgische procedures, evenals endoscopische onderzoeken, gebruiken medicijnen die ontspanning van skeletspieren veroorzaken (spierverslappers). Deze omvatten ditiline, een structureel analoog van acetylcholine. Het hecht zich aan H-cholinerge receptoren, maar in tegenstelling tot acetylcholine wordt het zeer langzaam vernietigd door het enzym acetylcholinesterase. Als gevolg van langdurige opening van ionenkanalen en aanhoudende depolarisatie van het membraan, wordt de geleiding van zenuwimpulsen verstoord en treedt spierontspanning op. Aanvankelijk werden deze eigenschappen gevonden in curare-gif, daarom worden dergelijke medicijnen genoemd curariform.
ONDERWERP 1.3. DENATURATIE VAN EIWITTEN EN DE MOGELIJKHEID VAN HUN SPONTANE RENATIVATIE
1. Aangezien de natieve conformatie van eiwitten behouden blijft als gevolg van zwakke interacties, veroorzaken veranderingen in de samenstelling en eigenschappen van de omgeving rond het eiwit, blootstelling aan chemische reagentia en fysieke factoren een verandering in hun conformatie (eigenschap van conformationele labiliteit). Het verbreken van een groot aantal bindingen leidt tot de vernietiging van de natieve conformatie en denaturatie van eiwitten.
Eiwitdenaturatie- dit is de vernietiging van hun natuurlijke conformatie onder de werking van denaturerende middelen, veroorzaakt door de breuk van zwakke bindingen die de ruimtelijke structuur van het eiwit stabiliseren. Denaturatie gaat gepaard met de vernietiging van de unieke driedimensionale structuur en het actieve centrum van het eiwit en het verlies van zijn biologische activiteit (Fig. 1.16).
Alle gedenatureerde moleculen van één eiwit krijgen een willekeurige conformatie die verschilt van andere moleculen van hetzelfde eiwit. De aminozuurradicalen die het actieve centrum vormen, zijn ruimtelijk ver van elkaar verwijderd, d.w.z. de specifieke bindingsplaats van het eiwit met het ligand wordt vernietigd. Tijdens denaturatie blijft de primaire structuur van eiwitten onveranderd.
Het gebruik van denaturerende middelen in biologisch onderzoek en geneeskunde. In biochemische onderzoeken worden, voordat verbindingen met een laag molecuulgewicht in biologisch materiaal worden bepaald, meestal eerst eiwitten uit de oplossing verwijderd. Voor dit doel wordt meestal trichloorazijnzuur (TCA) gebruikt. Na toevoeging van TCA aan de oplossing precipiteren gedenatureerde eiwitten en worden gemakkelijk verwijderd door filtratie (Tabel 1.1.)
In de geneeskunde worden denaturerende middelen vaak gebruikt om medische instrumenten en materialen in autoclaven te steriliseren (denatureringsmiddel - hoge temperatuur) en als antiseptica (alcohol, fenol, chlooramine) om besmette oppervlakken met pathogene microflora te behandelen.
2. Spontane eiwitvernieuwing- bewijs van het determinisme van de primaire structuur, conformatie en functie van eiwitten. Individuele eiwitten zijn producten van één gen die een identieke aminozuursequentie hebben en dezelfde conformatie in de cel verwerven. De fundamentele conclusie dat de primaire structuur van een eiwit al informatie over zijn conformatie en functie bevat, werd gemaakt op basis van het vermogen van sommige eiwitten (met name ribonuclease en myoglobine) om spontaan te regenereren - om hun oorspronkelijke conformatie te herstellen na denaturatie.
De vorming van de ruimtelijke structuren van een eiwit wordt uitgevoerd door de methode van zelfassemblage - een spontaan proces waarbij een polypeptideketen, die een unieke primaire structuur heeft, de neiging heeft een conformatie te accepteren met de laagste vrije energie in oplossing. Het vermogen om eiwitten te vernieuwen die hun primaire structuur behouden na denaturatie werd beschreven in een experiment met het enzym ribonuclease.
Ribonuclease is een enzym dat de bindingen tussen individuele nucleotiden in een RNA-molecuul afbreekt. Dit bolvormige eiwit heeft één polypeptideketen, waarvan de tertiaire structuur wordt gestabiliseerd door vele zwakke en vier disulfidebindingen.
De behandeling van ribonuclease met ureum, dat waterstofbruggen in het molecuul vernietigt, en een reductiemiddel dat disulfidebindingen verbreekt, leidt tot denaturatie van het enzym en het verlies van zijn activiteit.
Verwijdering van denaturerende middelen door dialyse resulteert in herstel van eiwitconformatie en -functie, d.w.z. aan de vernieuwing. (afb. 1.17).
 Rijst. 1.17. Denaturatie en renaissance van ribonuclease
Rijst. 1.17. Denaturatie en renaissance van ribonuclease
A - natieve conformatie van ribonuclease, in de tertiaire structuur waarvan er vier disulfidebindingen zijn; B - gedenatureerd ribonucleasemolecuul;
B - een nieuw leven ingeblazen ribonucleasemolecuul met verminderde structuur en functie
1. Volledige tabel 1.2.
Tabel 1.2. Classificatie van aminozuren door radicale polariteit
2. Schrijf de tetrapeptideformule:
Asp - Pro - Fen - Liz
a) isoleer in het peptide de herhalende groepen die de peptideruggengraat vormen en de variabele groepen weergegeven door aminozuurradicalen;
b) geef de N- en C-uiteinden aan;
c) onderstrepen peptidebindingen;
d) schrijf een ander peptide dat uit dezelfde aminozuren bestaat;
e) tel het aantal mogelijke varianten van een tetrapeptide met een vergelijkbare aminozuursamenstelling.
3. Verklaar de rol van de primaire structuur van eiwitten aan de hand van het voorbeeld van een vergelijkende analyse van twee structureel vergelijkbare en evolutionair nauw verwante peptidehormonen van de neurohypofyse van zoogdieren - oxytocine en vasopressine (Tabel 1.3).
Tabel 1.3. Structuur en functie van oxytocine en vasopressine
 Voor deze:
Voor deze:
a) vergelijk de samenstelling en aminozuursequentie van de twee peptiden;
b) zoek de overeenkomst van de primaire structuur van twee peptiden en de overeenkomst van hun biologische werking;
c) zoek de verschillen in de structuur van de twee peptiden en het verschil in hun functies;
d) een conclusie trekken over de invloed van de primaire structuur van peptiden op hun functies.
4. Beschrijf de belangrijkste stadia in de vorming van de conformatie van bolvormige eiwitten (secundaire, tertiaire structuren, het concept van een supersecundaire structuur). Geef de soorten bindingen aan die betrokken zijn bij de vorming van eiwitstructuren. Welke aminozuurradicalen kunnen deelnemen aan de vorming van hydrofobe interacties, ionische, waterstofbruggen.
Geef voorbeelden.
5. Geef een definitie van het begrip "conformationele labiliteit van eiwitten", geef de redenen voor het bestaan en de betekenis ervan aan.
6. Breid de betekenis van de volgende zin uit: "De werking van eiwitten is gebaseerd op hun specifieke interactie met het ligand", gebruik de termen en leg hun betekenis uit: eiwitconformatie, actief centrum, ligand, complementariteit, eiwitfunctie.
7. Leg met één voorbeeld uit wat domeinen zijn en wat hun rol in eiwitten is.
TAKEN VOOR ZELFCONTROLE
1. Correspondentie tot stand brengen.
Functionele groep in de aminozuurradicaal:
A. Carboxylgroep B. Hydroxylgroep C Guanidinegroep D. Thiolgroep D. Aminogroep
2. Kies de correcte antwoorden.
Aminozuren met polaire ongeladen radicalen zijn:
A. Cis B. Asn
B. Glu G. Drie
3. Kies de correcte antwoorden.
Aminozuurradicalen:
A. Geef de specificiteit van de primaire structuur B. Neem deel aan de vorming van de tertiaire structuur
B. Bevindt zich op het oppervlak van het eiwit, beïnvloedt de oplosbaarheid D. Vorm een actief centrum
E. Neem deel aan de vorming van peptidebindingen
4. Kies de correcte antwoorden.
Hydrofobe interacties kunnen ontstaan tussen aminozuurradicalen:
A. Tre Lei B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Kies de correcte antwoorden.
Ionische bindingen kunnen zich vormen tussen aminozuurradicalen:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gies Asp D. Asn april
6. Kies de correcte antwoorden.
Waterstofbindingen kunnen zich vormen tussen aminozuurradicalen:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Correspondentie tot stand brengen.
Het type binding dat betrokken is bij de vorming van de eiwitstructuur:
A. Primaire structuur B. Secundaire structuur
B. Tertiaire structuur
D. Supersecundaire structuur E. Conformatie.
1. Waterstofbindingen tussen de atomen van de peptideruggengraat
2. Zwakke bindingen tussen functionele groepen van aminozuurradicalen
3. Relaties tussen α-amino- en α-carboxylgroepen van aminozuren
8. Kies de correcte antwoorden. Trypsine:
A. Proteolytisch enzym B. Bevat twee domeinen
B. Hydrolyseert zetmeel
D. Het actieve centrum bevindt zich tussen de domeinen. D. Bestaat uit twee polypeptideketens.
9. Kies de correcte antwoorden. Atropine:
A. Neurotransmitter
B. Structurele analoog van acetylcholine
B. Interageert met H-cholinerge receptoren
D. Versterkt de geleiding van zenuwimpulsen door cholinerge synapsen
D. Competitieve remmer van M-cholinerge receptoren
10. Kies de juiste uitspraken. In eiwitten:
A. Primaire structuur bevat informatie over de structuur van zijn actieve centrum
B. Het actieve centrum wordt gevormd op het niveau van de primaire structuur
B. De conformatie is star gefixeerd door covalente bindingen
D. De actieve site kan interageren met een groep vergelijkbare liganden
vanwege de conformationele labiliteit van eiwitten D. Veranderingen in de omgeving kunnen de affiniteit van de actieve . beïnvloeden
centrum naar ligand
1.1-C, 2-D, 3-B.
3.A, B, C, D.
7.1-B, 2-D, 3-A.
8.A, B, C, D.
BASISVOORWAARDEN EN CONCEPTEN
1. Eiwit, polypeptide, aminozuren
2. Primaire, secundaire, tertiaire eiwitstructuur
3. Conformatie, natieve eiwitconformatie
4. Covalente en zwakke bindingen in een eiwit
5. conformatielabiliteit
6. Actief centrum van eiwitten
7. Liganden
8. Eiwitvouwing
9. Structurele analogen van liganden
10. Domein eiwitten
11. Eenvoudige en complexe eiwitten
12. Eiwitdenaturatie, denaturerende middelen
13. Renovatie van eiwitten
Taken oplossen
"Structurele organisatie van eiwitten en de basis van hun werking"
1. De belangrijkste functie van het eiwit, hemoglobine A (HbA), is het transporteren van zuurstof naar de weefsels. In de menselijke populatie zijn meerdere vormen van dit eiwit met veranderde eigenschappen en functies bekend - de zogenaamde abnormale hemoglobines. Er werd bijvoorbeeld gevonden dat hemoglobine S, gevonden in de erytrocyten van sikkelcelanemie (HbS)-patiënten, een lage oplosbaarheid heeft onder omstandigheden van lage partiële zuurstofdruk (zoals het geval is in veneus bloed). Dit leidt tot de vorming van aggregaten van dit eiwit. Het eiwit verliest zijn functie, slaat neer en rode bloedcellen krijgen een onregelmatige vorm (sommige vormen een sikkel) en worden sneller dan normaal vernietigd in de milt. Als gevolg hiervan ontwikkelt zich sikkelcelanemie.
Het enige verschil in de primaire structuur van HbA werd gevonden in het N-terminale gebied van de β-keten van hemoglobine. Vergelijk de N-terminale uiteinden van de β-keten en laat zien hoe veranderingen in de primaire structuur van een eiwit de eigenschappen en functies beïnvloeden.
 Voor deze:
Voor deze:
a) schrijf de aminozuurformules waarmee HBA verschilt en vergelijk de eigenschappen van deze aminozuren (polariteit, lading).
b) een conclusie trekken over de reden voor de afname van de oplosbaarheid en de verslechtering van het zuurstoftransport in het weefsel.
2. De figuur toont een diagram van de structuur van een eiwit dat een ligandbindingsplaats (actieve plaats) heeft. Leg uit waarom een eiwit selectief is in zijn ligandkeuze. Voor deze:
a) onthoud wat het actieve centrum van een eiwit is, en beschouw de structuur van het actieve centrum van het eiwit dat in de figuur wordt getoond;
b) schrijf de formules van de aminozuurradicalen die het actieve centrum vormen;
c) trek een ligand die specifiek zou kunnen interageren met het actieve centrum van het eiwit. Geef daarop de functionele groepen aan die in staat zijn bindingen te vormen met de aminozuurradicalen die het actieve centrum vormen;
d) de soorten bindingen aangeven die ontstaan tussen de ligand en de aminozuurradicalen van het actieve centrum;
e) leg uit waarop de specificiteit van de interactie van het eiwit met het ligand is gebaseerd.
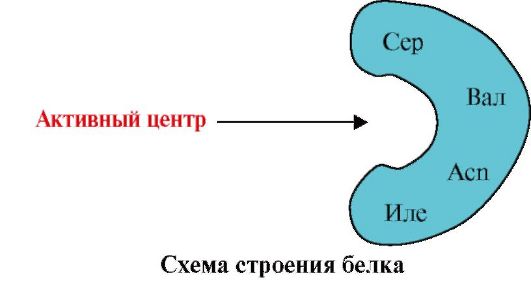 3.
De figuur toont de actieve plaats van het eiwit en enkele liganden.
3.
De figuur toont de actieve plaats van het eiwit en enkele liganden.
Bepaal welke van de liganden het meest waarschijnlijk een interactie aangaat met de actieve plaats van het eiwit en waarom.
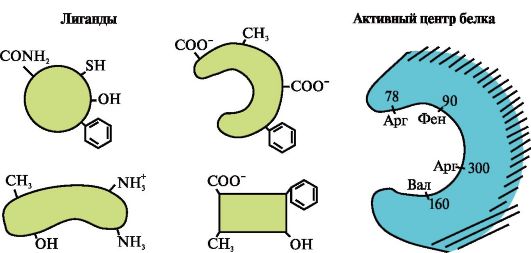 Welke soorten bindingen ontstaan tijdens de vorming van het eiwit-ligandcomplex?
Welke soorten bindingen ontstaan tijdens de vorming van het eiwit-ligandcomplex?
4. Structurele analogen van natuurlijke eiwitliganden kunnen worden gebruikt als geneesmiddelen om de activiteit van eiwitten te veranderen.
Acetylcholine is een zender van excitatie in neuromusculaire synapsen. Wanneer acetylcholine interageert met eiwitten - receptoren van het postsynaptische membraan van skeletspieren, openen ionkanalen en spiercontractie. Ditilin is een geneesmiddel dat bij sommige operaties wordt gebruikt om de spieren te ontspannen, omdat het de overdracht van zenuwimpulsen door neuromusculaire synapsen verstoort. Leg het werkingsmechanisme uit van ditiline als spierverslapper. Voor deze:
a) schrijf de formules van acetylcholine en ditiline en vergelijk hun structuren;
b) het mechanisme van de ontspannende werking van ditiline beschrijven.
5. Bij sommige ziekten stijgt de lichaamstemperatuur van de patiënt, wat wordt beschouwd als een beschermende reactie van het lichaam. Hoge temperaturen zijn echter schadelijk voor de eiwitten van het lichaam. Leg uit waarom bij temperaturen boven de 40°C de functie van eiwitten wordt verstoord en er een bedreiging voor het menselijk leven ontstaat. Om dit te doen, onthoud:
1) De structuur van eiwitten en bindingen die de structuur in de natieve conformatie houden;
2) Hoe verandert de structuur en functie van eiwitten bij toenemende temperatuur?;
3) Wat is homeostase en waarom is het belangrijk voor het behoud van de menselijke gezondheid.
Modulaire eenheid 2 OLIGOMERISCHE EIWITTEN ALS DOELSTELLINGEN VAN REGELGEVING. STRUCTURELE EN FUNCTIONELE EIWITVERSCHEIDENHEID. EIWITSCHEIDING EN ZUIVERINGSMETHODEN
Leerdoelen In staat zijn om:
1. Gebruik kennis over de kenmerken van de structuur en functies van oligomere eiwitten om de adaptieve mechanismen van regulering van hun functies te begrijpen.
2. De rol van chaperonnes in de synthese en instandhouding van eiwitconformatie onder celomstandigheden uitleggen.
3. Verklaar de verscheidenheid aan manifestaties van het leven door de verscheidenheid aan structuren en functies van eiwitten die in het lichaam worden gesynthetiseerd.
4. Analyse van de relatie tussen de structuur van eiwitten en hun functie met behulp van voorbeelden van vergelijking van verwante hemoproteïnen - myoglobine en hemoglobine, evenals vertegenwoordigers van vijf klassen van eiwitten van de immunoglobulinefamilie.
5. Toepassen van kennis over de eigenaardigheden van de fysisch-chemische eigenschappen van eiwitten om methoden te selecteren voor hun zuivering van andere eiwitten en onzuiverheden.
6. Interpreteer de resultaten van de kwantitatieve en kwalitatieve samenstelling van bloedplasma-eiwitten om de klinische diagnose te bevestigen of te verduidelijken.
Weten:
1. Kenmerken van de structuur van oligomere eiwitten en adaptieve mechanismen van regulering van hun functies naar het voorbeeld van hemoglobine.
2. De structuur en functie van chaperonnes en hun belang voor het in stand houden van de natieve conformatie van eiwitten onder celomstandigheden.
3. Principes van het combineren van eiwitten in families op basis van de gelijkenis van hun conformatie en functies op het voorbeeld van immunoglobulinen.
4. Methoden voor de scheiding van eiwitten op basis van de eigenaardigheden van hun fysisch-chemische eigenschappen.
5. Elektroforese van bloedplasma als methode om de kwalitatieve en kwantitatieve samenstelling van eiwitten te beoordelen.
ONDERWERP 1.4. KENMERKEN VAN DE STRUCTUUR EN WERKING VAN OLIGOMERISCHE EIWITTEN BIJ HET VOORBEELD VAN HEMOGLOBINE
1. Veel eiwitten bevatten meerdere polypeptideketens. Dergelijke eiwitten worden oligomeer, en individuele ketens - protomers. Protomeren in oligomere eiwitten zijn verbonden door vele zwakke niet-covalente bindingen (hydrofoob, ionisch, waterstof). Interactie
protomers wordt uitgevoerd dankzij: complementariteit hun contactvlakken.
Het aantal protomeren in oligomere eiwitten kan sterk variëren: hemoglobine bevat 4 protomeren, het aspartaataminotransferase-enzym bevat 12 protomeren en het tabaksmozaïekviruseiwit bevat 2.120 protomeren die zijn verbonden door niet-covalente bindingen. Bijgevolg kunnen oligomere eiwitten zeer hoge molecuulgewichten hebben.
De interactie van een protomeer met andere kan worden beschouwd als een speciaal geval van de interactie van een eiwit met een ligand, aangezien elke protomeer dient als een ligand voor andere protomeren. Het aantal en de methode van het combineren van protomeren in een eiwit wordt genoemd quaternaire eiwitstructuur.
Eiwitten kunnen protomeren met dezelfde of een verschillende structuur omvatten, homodimeren zijn bijvoorbeeld eiwitten die twee identieke protomeren bevatten en heterodimeren zijn eiwitten die twee verschillende protomeren bevatten.
Als eiwitten verschillende protomeren bevatten, kunnen daarop verschillende structuren van bindingsplaatsen met verschillende liganden worden gevormd. Wanneer het ligand aan het actieve centrum bindt, komt de functie van dit eiwit tot uiting. Het centrum dat zich op een ander protomeer bevindt, wordt allosterisch genoemd (anders dan het actieve). Contact opnemen een allosterische ligand of effector, hij presteert regulerende functie(afb. 1.18). De interactie van het allosterische centrum met de effector veroorzaakt conformationele veranderingen in de structuur van het gehele oligomere eiwit vanwege zijn conformationele labiliteit. Dit beïnvloedt de affiniteit van de actieve plaats voor een specifiek ligand en regelt de functie van dit eiwit. Een verandering in de conformatie en functie van alle protomeren bij interactie van een oligomeer eiwit met ten minste één ligand wordt coöperatieve conformatieveranderingen genoemd. Effectoren die de eiwitfunctie verbeteren, worden genoemd: activatoren, en de effectoren die de functie ervan remmen - remmers.
Zo hebben oligomere eiwitten, evenals eiwitten met een domeinstructuur, een nieuwe eigenschap in vergelijking met monomere eiwitten - het vermogen tot allosterische regulatie van functies (regulatie door de hechting van verschillende liganden aan het eiwit). Dit kan worden gezien door de structuren en functies van twee nauw verwante complexe eiwitten, myoglobine en hemoglobine, te vergelijken.
 Rijst. 1.18. Dimeer eiwitstructuurdiagram
Rijst. 1.18. Dimeer eiwitstructuurdiagram
2. De vorming van ruimtelijke structuren en het functioneren van myoglobine.
Myoglobine (MB) is een eiwit dat voorkomt in rode spieren en waarvan de belangrijkste functie het creëren van O2-reserves is, die nodig zijn voor intensieve spierarbeid. MB is een complex eiwit dat een eiwitdeel - apoMv - en een niet-eiwitdeel - heem bevat. De primaire structuur van apoMv bepaalt de compacte bolvormige conformatie en de structuur van de actieve plaats, waaraan het niet-eiwitdeel van myoglobine, heem, is gehecht. Zuurstof die uit het bloed naar de spieren komt, bindt zich aan Fe + 2 heem in myoglobine. MB is een monomeer eiwit met een zeer hoge affiniteit voor O 2, daarom vindt de afgifte van zuurstof door myoglobine alleen plaats tijdens intensieve spierarbeid, wanneer de partiële druk van O 2 sterk daalt.
Vorming van de MB conformatie. In rode spieren op ribosomen wordt tijdens translatie de primaire structuur van MB gesynthetiseerd, weergegeven door een specifieke sequentie van 153 aminozuurresiduen. De secundaire structuur MB bevat acht α-helices, de Latijnse letters van A tot H, waartussen zich niet-spiraalvormige secties bevinden. De tertiaire structuur van MB heeft de vorm van een compacte bol, in de verdieping waarvan tussen de F- en E -helices een actief centrum is (Fig. 1.19).
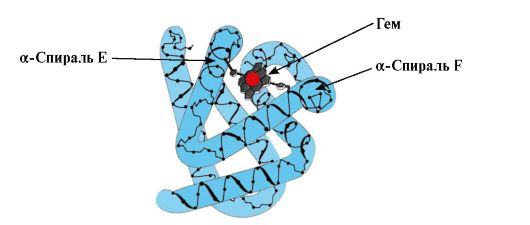 Rijst. 1.19. Myoglobine structuur
Rijst. 1.19. Myoglobine structuur
3. Kenmerken van de structuur en werking van het actieve centrum Мв. Het actieve centrum van MB wordt voornamelijk gevormd door hydrofobe aminozuurradicalen, die ver van elkaar verwijderd zijn in de primaire structuur (bijvoorbeeld Three 3 9 en Fen 138) Slecht in water oplosbare liganden - heem en O 2 - zijn gehecht aan het actieve centrum. Heme is een specifiek ligand van apoMv (Fig. 1.20), dat is gebaseerd op vier pyrroolringen verbonden door metenylbruggen; het Fe + 2-atoom bevindt zich in het midden, verbonden met de stikstofatomen van de pyrroolringen door vier coördinatiebindingen. In het actieve centrum van MB zijn er naast hydrofobe aminozuurradicalen ook residuen van twee aminozuren met hydrofiele radicalen - Gis E 7(Gis 64) en Zijn F 8(Gis 93) (afb. 1.21).
 Rijst. 1.20. De structuur van heem - het niet-eiwit deel van myoglobine en hemoglobine
Rijst. 1.20. De structuur van heem - het niet-eiwit deel van myoglobine en hemoglobine
 Rijst. 1.21. Locatie van heem en O2 in het actieve centrum van apomyoglobine en hemoglobine protomeren
Rijst. 1.21. Locatie van heem en O2 in het actieve centrum van apomyoglobine en hemoglobine protomeren
Heem is via een ijzeratoom covalent gebonden aan His F 8. O 2 hecht zich aan de klier aan de andere kant van het heemvlak. Zijn E 7 is nodig voor de juiste oriëntatie van O 2 en vergemakkelijkt de toevoeging van zuurstof aan Fe + 2 heem
Zijn F 8 vormt een coördinatiebinding met Fe + 2 en fixeert de heem stevig in het actieve centrum. Gis E 7 is nodig voor de juiste oriëntatie in het actieve centrum van een ander ligand, O 2, wanneer het interageert met Fe + 2 heem. De micro-omgeving van heem schept voorwaarden voor een sterke maar omkeerbare binding van O 2 met Fe + 2 en voorkomt dat water het hydrofobe actieve centrum binnendringt, wat kan leiden tot oxidatie tot Fe + 3.
De monomere structuur van MB en zijn actieve centrum bepaalt de hoge affiniteit van het eiwit voor O2.
4. Oligomere structuur van HB en regulatie van de affiniteit van HB tot 02-liganden. Menselijke hemoglobines- een familie van eiwitten, evenals myoglobine verwant aan complexe eiwitten (hemoproteïnen). Ze hebben een tetramere structuur en bevatten twee α-ketens, maar verschillen in de structuur van de andere twee polypeptideketens (2α-, 2x-ketens). De structuur van de tweede polypeptideketen bepaalt de kenmerken van het functioneren van deze vormen van HB. Ongeveer 98% van de hemoglobine van erytrocyten van een volwassene is hemoglobine A(2α-, 2p-ketens).
Tijdens de periode van intra-uteriene ontwikkeling zijn er twee hoofdtypen hemoglobines: embryonale HB(2α, 2ε), die wordt gevonden in de vroege stadia van de ontwikkeling van de foetus, en hemoglobine F (foetaal)- (2α, 2γ), dat de vroege foetale hemoglobine vervangt in de zesde maand van de intra-uteriene ontwikkeling en pas na de geboorte wordt vervangen door HB A.
HB A is een eiwit verwant aan myoglobine (MB) en wordt aangetroffen in de erytrocyten van een volwassene. De structuur van zijn individuele protomeren is vergelijkbaar met die van myoglobine. De secundaire en tertiaire structuren van myoglobine en hemoglobine protomeren lijken erg op elkaar, ondanks het feit dat in de primaire structuur van hun polypeptideketens slechts 24 aminozuurresiduen identiek zijn (de secundaire structuur van hemoglobine protomeren, zoals myoglobine, bevat acht a-helices, aangeduid met Latijnse letters van A tot H , en de tertiaire structuur ziet eruit als een compact bolletje). Maar in tegenstelling tot myoglobine heeft hemoglobine een oligomere structuur, bestaat het uit vier polypeptideketens die verbonden zijn door niet-covalente bindingen (Figuur 1.22).
Elke HB-protomeer is geassocieerd met een niet-eiwitdeel - heem en aangrenzende protomeren. De verbinding van het eiwitdeel van HB met heem is vergelijkbaar met die van myoglobine: in het actieve centrum van het eiwit zijn de hydrofobe delen van het heem omgeven door hydrofobe aminozuurradicalen, met uitzondering van His F 8 en His E 7 , die zich aan weerszijden van het heemvlak bevinden en een vergelijkbare rol spelen bij het functioneren van het eiwit en de binding ervan met zuurstof (zie de structuur van myoglobine).
 Rijst. 1.22. Oligomere structuur van hemoglobine
Rijst. 1.22. Oligomere structuur van hemoglobine
Bovendien, Gis E 7 vervult een belangrijke extra rol in het functioneren van de NV. Vrij heem heeft een 25.000 keer hogere affiniteit voor CO dan voor O2. CO wordt in kleine hoeveelheden in het lichaam gevormd en gezien de hoge affiniteit voor heem, zou het het transport van O 2 dat nodig is voor het leven van cellen kunnen verstoren. In de samenstelling van hemoglobine is de affiniteit van heem voor koolmonoxide echter slechts 200 keer groter dan de affiniteit voor O 2 vanwege de aanwezigheid van His E 7 in het actieve centrum. De rest van dit aminozuur creëert optimale omstandigheden voor heem om te binden aan O 2 en verzwakt de interactie van heem met CO.
5. De belangrijkste functie van HB is het transport van O 2 van de longen naar het weefsel. In tegenstelling tot monomeer myoglobine, dat een zeer hoge affiniteit voor O2 heeft en de functie vervult van het opslaan van zuurstof in rode spieren, biedt de oligomere structuur van hemoglobine:
1) snelle verzadiging van Hb met zuurstof in de longen;
2) het vermogen van HB om zuurstof te doneren in weefsels bij een relatief hoge partiële druk van 02 (20-40 mm Hg);
3) het vermogen om de affiniteit van HB tot O2 te reguleren.
6. Coöperatieve veranderingen in de conformatie van de hemoglobine-protomeren versnellen de binding van O 2 in de longen en de afgifte ervan in de weefsels. In de longen bevordert een hoge partiële druk van O 2 de binding aan HB in het actieve centrum van vier protomeren (2α en 2β). Het actieve centrum van elk protomeer, evenals in myoglobine, bevindt zich tussen twee a-helices (F en E) in een hydrofobe zak. Het bevat een niet-eiwitdeel - heem, gehecht aan het eiwitdeel door vele zwakke hydrofobe interacties en één sterke binding tussen Fe 2 + heem en His F 8 (zie Fig. 1.21).
In deoxyhemoglobine steekt door deze binding met His F 8 het Fe 2+-atoom uit het heemvlak naar histidine uit. Binding van O 2 met Fe 2 + vindt plaats aan de andere kant van het heem in het gebied van His E 7 met behulp van een enkele vrije coördinatiebinding. Gis E 7 biedt optimale omstandigheden voor de binding van O 2 aan heemijzer.
De aanhechting van O2 aan het Fe+2-atoom van één protomeer zorgt ervoor dat het naar het heem-vlak gaat, gevolgd door de histidinerest die eraan gebonden is.
 Rijst. 1.23. Verandering in de conformatie van het hemoglobine-protomeer bij combinatie met O 2
Rijst. 1.23. Verandering in de conformatie van het hemoglobine-protomeer bij combinatie met O 2
Dit leidt tot een verandering in de conformatie van alle polypeptideketens vanwege hun conformationele labiliteit. Het veranderen van de conformatie van andere ketens vergemakkelijkt hun interactie met de volgende O2-moleculen.
Het vierde O2-molecuul bindt 300 keer gemakkelijker aan hemoglobine dan het eerste (Fig. 1.24).
 Rijst. 1.24. Coöperatieve veranderingen in de conformatie van hemoglobine-protomeren tijdens de interactie met О2
Rijst. 1.24. Coöperatieve veranderingen in de conformatie van hemoglobine-protomeren tijdens de interactie met О2
In weefsels wordt elk volgend O2-molecuul gemakkelijker afgesplitst dan het vorige, ook door coöperatieve veranderingen in de conformatie van protomeren.
7. CO 2 en H +, gevormd tijdens het katabolisme van organische stoffen, verminderen de affiniteit van hemoglobine tot O 2 in verhouding tot hun concentratie. De energie die nodig is voor het functioneren van cellen wordt voornamelijk geproduceerd in de mitochondriën tijdens de oxidatie van organische stoffen met behulp van O 2 dat door hemoglobine uit de longen wordt geleverd. Als gevolg van de oxidatie van organische stoffen worden de eindproducten van hun verval gevormd: CO 2 en K 2 O, waarvan de hoeveelheid evenredig is met de intensiteit van de lopende oxidatieprocessen.
CO 2 diffusie van cellen naar het bloed en dringt door in de erytrocyten, waar het wordt omgezet in koolzuur onder invloed van het enzym carbanhydrase. Dit zwakke zuur dissocieert in een proton en bicarbonaat-ion.
H + kunnen zich bij de radicalen van His . voegen 14 6 in de α- en β-ketens van hemoglobine, d.w.z. in gebieden ver van de heem. Protonering van hemoglobine vermindert de affiniteit voor O 2, bevordert de eliminatie van O 2 uit oxyHb, de vorming van deoxyHb en verhoogt de toevoer van zuurstof naar de weefsels in verhouding tot het aantal gevormde protonen (Fig. 1.25).
Een toename van de hoeveelheid vrijgekomen zuurstof afhankelijk van een toename van de concentratie van H+ in erytrocyten wordt het Bohr-effect genoemd (naar de Deense fysioloog Christian Bohr, die dit effect voor het eerst ontdekte).
In de longen bevordert een hoge partiële zuurstofdruk de binding met deoxyHB, wat de affiniteit van het eiwit voor H+ vermindert. De vrijgekomen protonen onder de werking van koolzuuranhydrase interageren met bicarbonaten om СО 2 en Н 2 te vormen

 Rijst. 1.25. Afhankelijkheid van de Hb-affiniteit voor O 2 van de concentratie van CO 2 en protonen (Bohr-effect):
Rijst. 1.25. Afhankelijkheid van de Hb-affiniteit voor O 2 van de concentratie van CO 2 en protonen (Bohr-effect):
EEN- de invloed van de concentratie van CO 2 en H + op de afgifte van O 2 uit het complex met HB (Bohr-effect); B- oxygenatie van deoxyhemoglobine in de longen, vorming en afgifte van CO2.
Het resulterende CO 2 komt de alveolaire ruimte binnen en wordt verwijderd met uitgeademde lucht. Zo wordt de hoeveelheid zuurstof die vrijkomt door hemoglobine in weefsels gereguleerd door de producten van katabolisme van organische stoffen: hoe intensiever de afbraak van stoffen, bijvoorbeeld tijdens lichamelijke inspanning, hoe hoger de concentratie van CO 2 en H + en hoe meer zuurstof die de weefsels krijgen als gevolg van een afname van de affiniteit van Hb voor O2.
8. Allosterische regulatie van de Hb-affiniteit voor 02 door het ligand - 2,3-bis-fosfoglyceraat. In erytrocyten wordt een allosterische ligand van hemoglobine, 2,3-bisfosfoglyceraat (2,3-BFG), gesynthetiseerd uit het product van glucose-oxidatie, 1,3-bisfosfoglyceraat. Onder normale omstandigheden is de concentratie van 2,3-BFG hoog en vergelijkbaar met de concentratie van HB. 2,3-BFG heeft een sterke negatieve lading van -5.
 Bisfosfoglyceraat in weefselcapillairen, door te binden aan deoxyhemoglobine, verhoogt de afgifte van zuurstof aan het weefsel, waardoor de affiniteit van HB voor O2 afneemt.
Bisfosfoglyceraat in weefselcapillairen, door te binden aan deoxyhemoglobine, verhoogt de afgifte van zuurstof aan het weefsel, waardoor de affiniteit van HB voor O2 afneemt.
Er is een holte in het midden van het tetramere hemoglobinemolecuul. Het wordt gevormd door aminozuurresten van alle vier de protomeren (zie figuur 1.22). In weefselcapillairen leidt protonering van HB (Bohr-effect) tot een breuk in de binding tussen heemijzer en O2. In een molecuul
deoxyhemoglobine ontstaan, in vergelijking met oxyhemoglobine, extra ionische bindingen die protomeren verbinden, waardoor de afmetingen van de centrale holte toenemen in vergelijking met oxyhemoglobine. De centrale holte is de plaats waar 2,3-BFG zich hecht aan hemoglobine. Door het verschil in grootte van de centrale holte kan 2,3-BFG zich alleen hechten aan deoxyhemoglobine.
2,3-BFG interageert met hemoglobine in een gebied ver van de actieve centra van het eiwit en behoort tot allosterisch(regulerende) liganden, en de centrale holte van HB is allosterisch centrum. 2,3-BFG heeft een sterke negatieve lading en interageert met vijf positief geladen groepen van twee β-ketens van HB: de N-terminale α-aminogroep Val en de radicalen Lys 82 His 143 (Fig. 1.26).
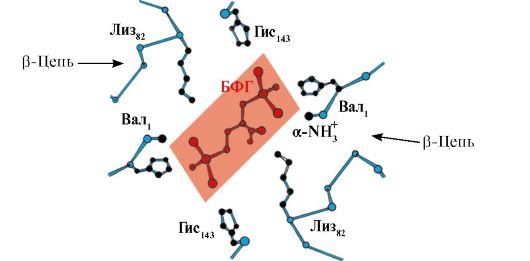 Rijst. 1.26. BPH in de centrale holte van deoxyhemoglobine
Rijst. 1.26. BPH in de centrale holte van deoxyhemoglobine
BPH bindt aan drie positief geladen groepen in elke β-keten.
In weefselcapillairen interageert het resulterende deoxyhemoglobine met 2,3-BFG en worden ionische bindingen gevormd tussen de positief geladen radicalen van de β-ketens en het negatief geladen ligand, die de conformatie van het eiwit veranderen en de affiniteit van HB voor O2. Een afname van de affiniteit van Hb voor O 2 draagt bij tot een efficiëntere afgifte van O 2 in het weefsel.
In de longen interageert zuurstof bij een hoge partiële druk met HB, waardoor het zich hecht aan het heemijzer; tegelijkertijd verandert de conformatie van het eiwit, neemt de centrale holte af en wordt 2,3-BFG verplaatst van het allosterische centrum
Zo hebben oligomere eiwitten nieuwe eigenschappen in vergelijking met monomere eiwitten. Aanhechting van liganden op locaties
ruimtelijk ver van elkaar (allosterisch), in staat conformationele veranderingen in het gehele eiwitmolecuul te veroorzaken. Door de interactie met regulerende liganden verandert de conformatie en past de functie van het eiwitmolecuul zich aan aan veranderingen in de omgeving.
ONDERWERP 1.5. ONDERHOUD VAN DE NATIEVE EIWITCONFORMATIE ONDER CELOMSTANDIGHEDEN
In cellen, tijdens de synthese van polypeptideketens, hun transport over membranen naar de overeenkomstige secties van de cel, tijdens vouwing (de vorming van een natieve conformatie) en tijdens de assemblage van oligomere eiwitten, evenals tijdens hun functioneren, intermediair, gevoelig tot aggregatie verschijnen onstabiele conformaties in de structuur van eiwitten. Hydrofobe radicalen, in de natieve conformatie die gewoonlijk verborgen zijn in een eiwitmolecuul, bevinden zich aan de oppervlakte in een onstabiele conformatie en hebben de neiging om te combineren met dezelfde slecht in water oplosbare groepen van andere eiwitten. In de cellen van alle bekende organismen zijn speciale eiwitten gevonden die zorgen voor een optimale vouwing van celeiwitten, hun natieve conformatie stabiliseren tijdens het functioneren en, belangrijker nog, de structuur en functie van intracellulaire eiwitten behouden in geval van homeostase-verstoring. Deze eiwitten heten "begeleiders", wat in het Frans "oppas" betekent.
1. Moleculaire chaperonnes en hun rol bij het voorkomen van eiwitdenaturatie.
Chaperones (III) worden geclassificeerd op basis van de massa van de subeenheid. Chaperones met een hoog molecuulgewicht hebben een massa van 60 tot 110 kDa. Onder hen zijn drie klassen het meest bestudeerd: Sh-60, Sh-70 en Sh-90. Elke klasse omvat een familie van verwante eiwitten. De samenstelling van III-70 omvat dus eiwitten met een molecuulgewicht van 66 tot 78 kDa. Chaperones met een laag molecuulgewicht hebben een molecuulgewicht van 40 tot 15 kDa.
Onder de chaperonnes worden onderscheiden constitutief eiwitten, waarvan de hoge basale synthese niet afhankelijk is van stresseffecten op de lichaamscellen, en induceerbaar, waarvan de synthese onder normale omstandigheden zwak is, maar sterk toeneemt onder stress. Induceerbare chaperonnes worden ook wel "heatshock-eiwitten" genoemd omdat ze voor het eerst werden ontdekt in cellen die aan hoge temperaturen werden blootgesteld. In cellen is door de hoge concentratie aan eiwitten de spontane vernieuwing van gedeeltelijk gedenatureerde eiwitten moeilijk. III-70 kan het beginnende proces van denaturatie voorkomen en de natieve conformatie van eiwitten helpen herstellen. Moleculaire chaperonnes-70- een sterk geconserveerde klasse van eiwitten die in alle delen van de cel wordt aangetroffen: cytoplasma, kern, endoplasmatisch reticulum, mitochondriën. Aan het carboxyluiteinde van de enige III-70-polypeptideketen bevindt zich een gebied dat een groef is die in staat is om te interageren met peptiden van lengte
van 7 tot 9 aminozuurresten verrijkt met hydrofobe radicalen. Dergelijke gebieden in bolvormige eiwitten komen ongeveer elke 16 aminozuren voor. III-70 zijn in staat eiwitten te beschermen tegen temperatuurinactivatie en de conformatie en activiteit van gedeeltelijk gedenatureerde eiwitten te herstellen.
2. De rol van chaperonnes bij het vouwen van eiwitten. Wanneer eiwitten op het ribosoom worden gesynthetiseerd, wordt het N-terminale gebied van het polypeptide eerder gesynthetiseerd dan het C-terminale gebied. De volledige aminozuursequentie van het eiwit is vereist om de natieve conformatie te vormen. Tijdens het proces van eiwitsynthese zijn chaperones-70, vanwege de structuur van hun actieve centrum, in staat om de aggregatiegevoelige gebieden van het polypeptide verrijkt met hydrofobe aminozuurradicalen te sluiten totdat de synthese voltooid is (Figuur 1.27, A).
 Rijst. 1.27. Deelname van chaperonnes aan eiwitvouwing
Rijst. 1.27. Deelname van chaperonnes aan eiwitvouwing
A - deelname van chaperonnes-70 aan de preventie van hydrofobe interacties tussen regio's van het gesynthetiseerde polypeptide; B - vorming van natieve eiwitconformatie in het chaperonnecomplex
Veel eiwitten met een hoog molecuulgewicht met een complexe conformatie, bijvoorbeeld een domeinstructuur, vouwen in een speciale ruimte gevormd door III-60. Sh-60 functioneren als een oligomeer complex bestaande uit 14 subeenheden. Ze vormen twee holle ringen die elk uit zeven subeenheden bestaan, deze ringen zijn met elkaar verbonden. Elke III-60-subeenheid bestaat uit drie domeinen: apicaal (apical), verrijkt met hydrofobe radicalen die naar de ringholte zijn gericht, intermediair en equatoriaal (Fig. 1.28).
 Rijst. 1.28. De structuur van het chaperonine-complex, bestaande uit 14 Sh-60
Rijst. 1.28. De structuur van het chaperonine-complex, bestaande uit 14 Sh-60
A - zijaanzicht; B - bovenaanzicht
De gesynthetiseerde eiwitten, die elementen op het oppervlak hebben die kenmerkend zijn voor ongevouwen moleculen, in het bijzonder hydrofobe radicalen, komen de holte van de chaperonneringen binnen. In de specifieke omgeving van deze holtes vindt een opsomming van mogelijke conformaties plaats totdat de enige, energetisch meest gunstige, is gevonden (Fig. 1.27, B). De vorming van conformaties en de afgifte van eiwit gaat gepaard met hydrolyse van ATP in het equatoriale gebied. Typisch vereist een dergelijke chaperonne-afhankelijke vouwing een aanzienlijke hoeveelheid energie.
Naast deelname aan de vorming van de driedimensionale structuur van eiwitten en de heropleving van gedeeltelijk gedenatureerde eiwitten, zijn chaperonnes ook vereist voor fundamentele processen zoals de assemblage van oligomere eiwitten, herkenning en transport van gedenatureerde eiwitten naar lysosomen, transport van eiwitten over membranen, en deelname aan de regulatie van de activiteit van eiwitcomplexen.
ONDERWERP 1.6. VERSCHEIDENHEID VAN EIWITTEN. EIWITFAMILIES VOLGENS HET VOORBEELD VAN IMMUNOGLOBULINEN
1. Eiwitten spelen een beslissende rol in het leven van individuele cellen en het hele meercellige organisme, en hun functies zijn verrassend divers. Dit wordt bepaald door de kenmerken van de primaire structuur en conformaties van eiwitten, de uniciteit van de structuur van het actieve centrum en het vermogen om specifieke liganden te binden.
Slechts een zeer kleine fractie van alle mogelijke varianten van peptideketens kan een stabiele ruimtelijke structuur aannemen; meerderheid
van hen kunnen veel conformaties aannemen met ongeveer dezelfde Gibbs-energie, maar met verschillende eigenschappen... De primaire structuur van de meeste bekende eiwitten die door biologische evolutie zijn geselecteerd, zorgt voor een uitzonderlijke stabiliteit van een van de conformaties die de eigenaardigheden van het functioneren van dit eiwit bepaalt.
2. Families van eiwitten. Binnen dezelfde biologische soort kunnen substituties van aminozuurresiduen leiden tot het ontstaan van verschillende eiwitten die verwante functies vervullen en homologe aminozuursequenties hebben. Dergelijke verwante eiwitten hebben opvallend vergelijkbare conformaties: het aantal en de tussenplaatsing van a-helices en/of β-structuren, de meeste bochten en bochten van de polypeptideketens zijn vergelijkbaar of identiek. Eiwitten met homologe regio's van de polypeptideketen, vergelijkbare conformatie en verwante functies worden geïsoleerd in eiwitfamilies. Voorbeelden van eiwitfamilies: serineproteïnasen, immunoglobulinefamilie, myoglobinefamilie.
Serine Proteïnasen- een familie van eiwitten die de functie van proteolytische enzymen vervullen. Deze omvatten spijsverteringsenzymen - chymotrypsine, trypsine, elastase en vele bloedstollingsfactoren. Deze eiwitten hebben identieke aminozuren op 40% van de posities en een zeer nauwe conformatie (Fig. 1.29).

Rijst. 1.29. Ruimtelijke structuren van elastase (A) en chymotrypsine (B)
Sommige aminozuursubstituties hebben geleid tot een verandering in de substraatspecificiteit van deze eiwitten en het ontstaan van functionele diversiteit binnen de familie.
3. De familie van immunoglobulinen. Eiwitten van de immunoglobuline-superfamilie, die drie families van eiwitten omvat, spelen een grote rol in het functioneren van het immuunsysteem:
Antilichamen (immunoglobulinen);
T-lymfocytreceptoren;
Eiwitten van het belangrijkste histocompatibiliteitscomplex - MHC-klassen 1 en 2 (Major Histocompatibiliteitscomplex).
Al deze eiwitten hebben een domeinstructuur, bestaan uit homologe immuunachtige domeinen en vervullen vergelijkbare functies: ze interageren met vreemde structuren, hetzij opgelost in het bloed, lymfe of intercellulaire vloeistof (antilichamen), of gelokaliseerd op het oppervlak van cellen (eigen of buitenlands).
4. Antilichamen- specifieke eiwitten geproduceerd door B-lymfocyten als reactie op het binnendringen van een vreemde structuur in het lichaam, genaamd antigeen.
Kenmerken van de structuur van antilichamen
De eenvoudigste antilichaammoleculen bestaan uit vier polypeptideketens: twee identieke lichte - L, die ongeveer 220 aminozuren bevat, en twee identieke zware - H, bestaande uit 440-700 aminozuren. Alle vier de ketens in een antilichaammolecuul zijn verbonden door vele niet-covalente bindingen en vier disulfidebindingen (Fig. 1.30).
Lichte antilichaamketens bestaan uit twee domeinen: variabel (VL), gelokaliseerd in het N-terminale gebied van de polypeptideketen, en constant (CL), gelokaliseerd op het C-uiteinde. Zware ketens hebben meestal vier domeinen: één variabele (VH) aan het N-uiteinde en drie constante (CH1, CH2, CH3) (zie Fig. 1.30). Elk immunoglobulinedomein heeft een β-sheet-superstructuur waarin twee cysteïneresiduen zijn verbonden door een disulfidebinding.
Tussen de twee constante domeinen CH1 en CH2 bevindt zich een gebied met een groot aantal prolineresiduen, die de vorming van een secundaire structuur en de interactie van naburige H-ketens in dit segment voorkomen. Dit scharniergebied verschaft flexibiliteit aan het antilichaammolecuul. Tussen de variabele domeinen van de zware en lichte ketens bevinden zich twee identieke antigeenbindingsplaatsen (actieve plaatsen voor antigeenbinding), daarom worden dergelijke antilichamen vaak bivalenten. Niet de gehele aminozuursequentie van de variabele gebieden van beide ketens is betrokken bij de binding van het antigeen aan het antilichaam, maar slechts 20-30 aminozuren die zich in de hypervariabele gebieden van elke keten bevinden. Het zijn deze regio's die het unieke vermogen van elk type antilichaam om een interactie aan te gaan met het overeenkomstige complementaire antigeen bepalen.
Antilichamen zijn een van de verdedigingslinies van het lichaam tegen binnendringende buitenaardse organismen. Hun werking kan in twee fasen worden verdeeld: de eerste fase is de herkenning en binding van het antigeen op het oppervlak van vreemde organismen, wat mogelijk is door de aanwezigheid van antigeenbindingsplaatsen in de structuur van het antilichaam; de tweede fase is de start van het proces van inactivatie en vernietiging van het antigeen. De specificiteit van de tweede fase hangt af van de klasse van antilichamen. Er zijn vijf klassen van zware ketens, die van elkaar verschillen in de structuur van constante domeinen: α, δ, ε, γ en μ, volgens welke vijf klassen van immunoglobulinen worden onderscheiden: A, D, E, G en M.
Structurele kenmerken van zware ketens geven de scharniergebieden en C-terminale gebieden van zware ketens een conformatiekenmerk van elke klasse. Na binding van het antigeen aan het antilichaam bepalen conformationele veranderingen in de constante domeinen de route voor antigeenverwijdering.
 Rijst. 1.30 Domeinstructuur van IgG
Rijst. 1.30 Domeinstructuur van IgG
Immunoglobulinen M
Immunoglobulinen M hebben twee vormen.
Monomere vorm- 1e klasse antilichamen geproduceerd door de zich ontwikkelende B-lymfocyt. Vervolgens schakelen veel B-cellen over op de productie van andere klassen antilichamen, maar met dezelfde antigeenbindingsplaats. IgM is opgenomen in het membraan en werkt als een antigeenherkenningsreceptor. De opname van IgM in het celmembraan is mogelijk door de aanwezigheid van 25 hydrofobe aminozuurresiduen in het staartgedeelte van het gebied.
Secretoire vorm van IgM bevat vijf monomere subeenheden die aan elkaar zijn gekoppeld door disulfidebindingen en een extra J-keten van het polypeptide (Fig. 1.31). Zware ketens van monomeren van deze vorm bevatten geen hydrofobe staart. De pentameer heeft 10 antigeenbindingsplaatsen en is daarom effectief in het herkennen en verwijderen van antigeen dat voor het eerst in het lichaam is binnengekomen. De secretoire vorm van IgM is de belangrijkste klasse van antilichamen die tijdens de primaire immuunrespons in de bloedbaan worden uitgescheiden. Binding van IgM aan een antigeen verandert de conformatie van IgM en induceert zijn binding aan de eerste eiwitcomponent van het complementsysteem (het complementsysteem is een set eiwitten die betrokken zijn bij de vernietiging van het antigeen) en activeert dit systeem. Als het antigeen zich op het oppervlak van het micro-organisme bevindt, veroorzaakt het complementsysteem een schending van de integriteit van het celmembraan en de dood van de bacteriële cel.
Immunoglobulinen G
In kwantitatieve termen overheerst deze klasse van immunoglobulinen in het bloed (75% van alle Ig). IgG - monomeren, de belangrijkste klasse van antilichamen, uitgescheiden in de bloedbaan tijdens een secundaire immuunrespons. Na de interactie van IgG met de oppervlakte-antigenen van micro-organismen, kan het antigeen-antilichaamcomplex eiwitten van het complementsysteem binden en activeren of kan het een interactie aangaan met specifieke receptoren van macrofagen en neutrofielen. Interactie met fagocyten leidt
 Rijst. 1.31. De structuur van de secretoire vorm van IgM
Rijst. 1.31. De structuur van de secretoire vorm van IgM
tot de absorptie van antigeen-antilichaamcomplexen en hun vernietiging in fagosomen van cellen. IgG is de enige klasse van antilichamen die de placentabarrière kunnen passeren en de foetus intra-uteriene bescherming bieden tegen infecties.
Immunoglobulinen A
De belangrijkste klasse van antilichamen die aanwezig zijn in secreties (melk, speeksel, respiratoire en intestinale secreties). IgA wordt voornamelijk in dimere vorm uitgescheiden, waarbij de monomeren aan elkaar zijn gekoppeld via een extra J-keten (Fig. 1.32).
IgA heeft geen interactie met het complementsysteem en fagocytische cellen, maar door te binden aan micro-organismen, voorkomen antilichamen dat ze zich hechten aan epitheelcellen en het lichaam binnendringen.
Immunoglobulinen E
Immunoglobulinen E worden weergegeven door monomeren waarin de zware -ketens, zoals de μ-ketens van immunoglobulinen M, één variabele en vier constante domeinen bevatten. IgE, na secretie, binden
 Rijst. 1.32. IgA-structuur
Rijst. 1.32. IgA-structuur
C-terminale plaatsen met overeenkomstige receptoren op het oppervlak van mestcellen en basofielen. Als resultaat worden ze receptoren voor antigenen op het oppervlak van deze cellen (Fig. 1.33).
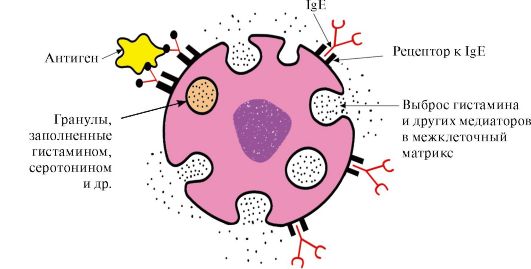 Rijst. 1.33. Interactie van IgE met antigeen op het mestceloppervlak
Rijst. 1.33. Interactie van IgE met antigeen op het mestceloppervlak
Nadat het antigeen zich hecht aan de overeenkomstige antigeenbindingsplaatsen van IgE, ontvangen de cellen een signaal voor de afscheiding van biologisch actieve stoffen (histamine, serotonine), die grotendeels verantwoordelijk zijn voor de ontwikkeling van de ontstekingsreactie en voor de manifestatie van allergische reacties zoals astma, urticaria, hooikoorts.
Immunoglobulinen D
Immunoglobulinen D worden in zeer kleine hoeveelheden in serum aangetroffen; het zijn monomeren. Zware δ-ketens hebben één variabele en drie constante domeinen. IgD werkt als receptor voor B-lymfocyten, andere functies zijn nog onbekend. De interactie van specifieke antigenen met receptoren op het oppervlak van B-lymfocyten (IgD) leidt tot de overdracht van deze signalen naar de cel en de activering van mechanismen die zorgen voor de vermenigvuldiging van deze kloon van lymfocyten.
ONDERWERP 1.7. FYSISCHE EN CHEMISCHE EIGENSCHAPPEN VAN EIWITTEN EN METHODEN VAN HUN SCHEIDING
1. Individuele eiwitten verschillen in hun fysische en chemische eigenschappen:
De vorm van de moleculen;
Moleculair gewicht;
De totale lading, waarvan de waarde afhangt van de verhouding van de anionische en kationische groepen aminozuren;
De verhouding van polaire en niet-polaire aminozuurradicalen op het oppervlak van moleculen;
De mate van resistentie tegen de effecten van verschillende denaturerende middelen.
2. De oplosbaarheid van eiwitten is afhankelijk van op de eigenschappen van de hierboven genoemde eiwitten, evenals op de samenstelling van het medium waarin het eiwit is opgelost (pH-waarden, zout samenstelling, temperatuur, de aanwezigheid van andere organische stoffen die kunnen interageren met het eiwit). De grootte van de lading van eiwitmoleculen is een van de factoren die hun oplosbaarheid beïnvloeden. Met een verlies van lading op het iso-elektrische punt, aggregeren eiwitten gemakkelijker en precipiteren. Dit geldt met name voor gedenatureerde eiwitten, die hydrofobe aminozuurradicalen op hun oppervlak hebben.
Op het oppervlak van een eiwitmolecuul bevinden zich zowel positief als negatief geladen aminozuurradicalen. Het aantal van deze groepen, en dus de totale lading van eiwitten, hangt af van de pH van het medium, d.w.z. de verhouding van de concentratie van H + - en OH - -groepen. In een zure omgeving een verhoging van de concentratie van H + leidt tot onderdrukking van de dissociatie van carboxylgroepen -COO - + H +> - COOH en een afname van de negatieve lading van eiwitten. In een alkalisch medium leidt de binding van overtollig OH - door protonen gevormd tijdens de dissociatie van aminogroepen -NH 3 + + OH - - NH 2 + H 2 O met de vorming van water tot een afname van de positieve lading van eiwitten. De pH-waarde waarbij het eiwit totaal nul is, heet iso-elektrisch punt (IEP). In IEP is het aantal positief en negatief geladen groepen hetzelfde, d.w.z. het eiwit is in een iso-elektrische toestand.
3. Scheiding van individuele eiwitten. Kenmerken van de structuur en werking van het lichaam zijn afhankelijk van de set eiwitten die erin wordt gesynthetiseerd. De studie van de structuur en eigenschappen van eiwitten is onmogelijk zonder hun isolatie van de cel en zuivering van andere eiwitten en organische moleculen. Stadia van isolatie en zuivering van individuele eiwitten:
cel vernietiging van het bestudeerde weefsel en het verkrijgen van een homogenaat.
Scheiding van het homogenaat in fracties door centrifugatie, het verkrijgen van een nucleaire, mitochondriale, cytosolische of andere fractie die het gewenste eiwit bevat.
Selectieve warmtedenaturatie- kortstondige verhitting van de eiwitoplossing, waarbij het mogelijk is een deel van de gedenatureerde eiwitonzuiverheden te verwijderen (in het geval dat het eiwit relatief thermostabiel is).
Uitzouten. Verschillende eiwitten precipiteren bij verschillende zoutconcentraties in oplossing. Door de zoutconcentratie geleidelijk te verhogen, is het mogelijk om een aantal afzonderlijke fracties te verkrijgen met een overwegend gehalte aan het uitgescheiden eiwit in een ervan. Ammoniumsulfaat wordt het meest gebruikt voor eiwitfractionering. Eiwitten met de laagste oplosbaarheid slaan neer bij lage zoutconcentraties.
Gelfiltratie- een methode om moleculen te zeven door gezwollen Sephadex-korrels (driedimensionale polysacharide dextranketens met poriën). De snelheid waarmee eiwitten door een met Sephadex gevulde kolom gaan, hangt af van hun molecuulgewicht: hoe kleiner de massa van eiwitmoleculen, hoe gemakkelijker ze in de korrels doordringen en daar langer blijven, hoe groter de massa, hoe sneller ze uit de kolom.
Ultracentrifugatie- een methode waarbij eiwitten in een centrifugebuis in een ultracentrifugerotor worden geplaatst. Wanneer de rotor draait, is de sedimentatiesnelheid van eiwitten evenredig met hun molecuulgewicht: fracties van zwaardere eiwitten bevinden zich dichter bij de bodem van de reageerbuis, lichtere bevinden zich dichter bij het oppervlak.
elektroforese- een methode gebaseerd op verschillen in de bewegingssnelheid van eiwitten in een elektrisch veld. Deze waarde is evenredig met de lading van eiwitten. Eiwitelektroforese wordt uitgevoerd op papier (in dit geval is de bewegingssnelheid van eiwitten alleen evenredig met hun lading) of in polyacrylamidegel met een bepaalde poriegrootte (de bewegingssnelheid van eiwitten is evenredig met hun lading en molecuulgewicht) .
Ionenuitwisselingschromatografie- een fractioneringsmethode gebaseerd op de binding van geïoniseerde groepen van eiwitten met tegengesteld geladen groepen van ionenuitwisselingsharsen (onoplosbare polymere materialen). De sterkte van de binding van het eiwit aan de hars is evenredig met de lading van het eiwit. Eiwitten die zijn geadsorbeerd aan een ionenuitwisselingspolymeer kunnen met toenemende concentraties worden afgewassen met NaCl-oplossingen; hoe lager de lading van het eiwit, hoe lager de concentratie van NaCl nodig zal zijn om het eiwit dat aan de ionische groepen van de hars is gebonden, af te wassen.
Affiniteitschromatografie- de meest specifieke methode om individuele eiwitten te isoleren Een ligand van een eiwit wordt covalent gebonden aan een inert polymeer. Wanneer een oplossing van eiwitten door een kolom met een polymeer wordt geleid vanwege de complementaire binding van het eiwit aan het ligand, wordt alleen het eiwit dat specifiek is voor het gegeven ligand op de kolom geadsorbeerd.
Dialyse- een methode die wordt gebruikt om verbindingen met een laag molecuulgewicht uit een oplossing van het geïsoleerde eiwit te verwijderen. De methode is gebaseerd op het onvermogen van eiwitten om door een semipermeabel membraan te gaan, in tegenstelling tot stoffen met een laag molecuulgewicht. Het wordt gebruikt voor de zuivering van eiwitten uit onzuiverheden met een laag molecuulgewicht, bijvoorbeeld uit zouten na uitzouten.
DOELSTELLINGEN VOOR OUT-OF-CONTROLE WERKEN
1. Vul de tabel in. 1.4.
Tabel 1.4. Vergelijkende analyse van de structuur en functies van verwante eiwitten - myoglobine en hemoglobine
a) onthoud de structuur van het actieve centrum MB en Hb. Wat is de rol van hydrofobe aminozuurradicalen bij de vorming van actieve centra van deze eiwitten? Beschrijf de structuur van het MB- en Hb-actieve centrum en de mechanismen van ligandaanhechting eraan. Wat is de rol van His F 8- en His E 7-residuen in het functioneren van het actieve centrum MB en Hv?
b) welke nieuwe eigenschappen heeft een nauw verwant oligomeer eiwit, hemoglobine, in vergelijking met monomeer myoglobine? Leg de rol uit van coöperatieve veranderingen in de conformatie van protomeren in het hemoglobinemolecuul, het effect van CO2- en protonconcentraties op de zuurstofaffiniteit van hemoglobine, evenals de rol van 2,3-BPH in de allosterische regulatie van de Hb-functie.
2. Karakteriseren van moleculaire chaperonnes, met aandacht voor de relatie tussen hun structuur en functie.
3. Welke eiwitten zijn onderverdeeld in families? Bepaal met behulp van de immunoglobulinefamilie als voorbeeld de vergelijkbare structurele kenmerken en verwante functies van de eiwitten van deze familie.
4. Gezuiverde individuele eiwitten zijn vaak nodig voor biochemische en medische doeleinden. Leg uit welke fysisch-chemische eigenschappen van eiwitten de gebruikte methoden zijn voor hun scheiding en zuivering.
TAKEN VOOR ZELFCONTROLE
1. Kies de correcte antwoorden.
Hemoglobine functies:
A. Transport van O 2 van de longen naar de weefsels B. Transport van H+ van de weefsels naar de longen
B. Handhaving van een constante bloed-pH D. Transport van CO 2 van de longen naar het weefsel
E. Transport van CO 2 van weefsels naar longen
2. Kies de correcte antwoorden. Ligandα -protomeer HB is: A. Heme
B. Zuurstof
B. CO G. 2,3-BFG
E. β-Protomeer
3. Kies de correcte antwoorden.
Hemoglobine in tegenstelling tot myoglobine:
A. Heeft een quaternaire structuur
B. Secundaire structuur wordt alleen weergegeven door -helices
B. Verwijst naar complexe eiwitten
D. Interageert met een allosterisch ligand D. Covalent gebonden aan heem
4. Kies de correcte antwoorden.
De affiniteit van Нв voor О 2 neemt af:
A. Met de toevoeging van één O2-molecuul B. Met de eliminatie van één O2-molecuul
B. Bij interactie met 2,3-BFG
D. Indien gehecht aan protomeren H + D. Met een afname van de concentratie van 2,3-BFG
5. Correspondentie tot stand brengen.
Typisch voor HB-types:
A. Vormt in deoxyvorm fibrillaire aggregaten B. Bevat twee α- en twee δ-ketens
B. De overheersende vorm van HB in de erytrocyten van een volwassene D. In het actieve centrum bevat heem met Fe + 3
E. Bevat twee α- en twee γ-ketens 1. НвА 2.
6. Correspondentie tot stand brengen.
Liganden HB:
A. Bindt aan HB in het allosterische centrum
B. Heeft een zeer hoge affiniteit voor het actieve centrum HB
B. Samenvoegen, verhoogt de affiniteit van Hb voor O 2 G. Oxideert Fe + 2 tot Fe + 3
E. Vormt een covalente binding met hisF8
7. Kies de correcte antwoorden.
Begeleiders:
A. Eiwitten aanwezig in alle delen van de cel
B. Synthese wordt versterkt door stress
B. Deelnemen aan de hydrolyse van gedenatureerde eiwitten
D. Neem deel aan het in stand houden van de natieve conformatie van eiwitten
E. Creëer organellen waarin eiwitconformatie wordt gevormd
8. Stel de correspondentie in. Immunoglobulinen:
A. De secretoire vorm is pentameric
B. Klasse Ig, penetreert de placentabarrière
B. Ig - mestcelreceptor
D. De belangrijkste klasse van Ig, aanwezig in de secreties van epitheelcellen. E. B-lymfocytreceptor, waarvan de activering zorgt voor celproliferatie
9. Kies de correcte antwoorden.
Immunoglobulinen E:
A. Geproduceerd door macrofagen B. Hebben zware ε-ketens.
B. Zijn opgenomen in het membraan van T-lymfocyten
D. Speel de rol van membraanreceptoren voor antigenen op mestcellen en basofielen
E. Verantwoordelijk voor de manifestatie van allergische reacties
10. Kies de correcte antwoorden.
De methode voor het scheiden van eiwitten is gebaseerd op verschillen in hun molecuulgewicht:
A. Gelfiltratie
B. Ultracentrifugatie
B. Polyacrylamidegelelektroforese D. Ionenuitwisselingschromatografie
D. Affiniteitschromatografie
11. Kies het juiste antwoord.
De methode voor het scheiden van eiwitten is gebaseerd op verschillen in oplosbaarheid in water:
A. Gelfiltratie B. Uitzouten
B. Ionenuitwisselingschromatografie D. Affiniteitschromatografie
D. Polyacrylamidegelelektroforese
STANDAARDEN VAN ANTWOORDEN OP "TAKEN VOOR ZELFCONTROLE"
1.A, B, C, D
2.A, B, C, D
5.1-B, 2-A, 3-D
6.1-B, 2-B, 3-A
7.A, B, D, D
8.1-D; 2-B, 3-C
BASISVOORWAARDEN EN CONCEPTEN
1. Oligomere eiwitten, protomeer, quaternaire structuur van eiwitten
2. Coöperatieve veranderingen in de conformatie van protomeren
3. Bohr-effect
4. Allosterische regulatie van eiwitfuncties, allosterisch centrum en allosterische effector
5. Moleculaire chaperonnes, heat shock-eiwitten
6. Eiwitfamilies (serineproteasen, immunoglobulinen)
7. IgM-, G-, E-, A-verbinding van structuur met functie
8. Totale lading van eiwitten, iso-elektrisch punt van eiwitten
9. Elektroforese
10. Uitzouten
11. Gelfiltratie
12. Ionenuitwisselingschromatografie
13. Ultracentrifugatie
14. Affiniteitschromatografie
15. Elektroforese van bloedplasma-eiwitten
TAKEN VOOR CONTROLEWERKEN
1. Vergelijk de afhankelijkheid van de mate van verzadiging van hemoglobine (Hb) en myoglobine (Mb) met zuurstof op de partiële druk in de weefsels
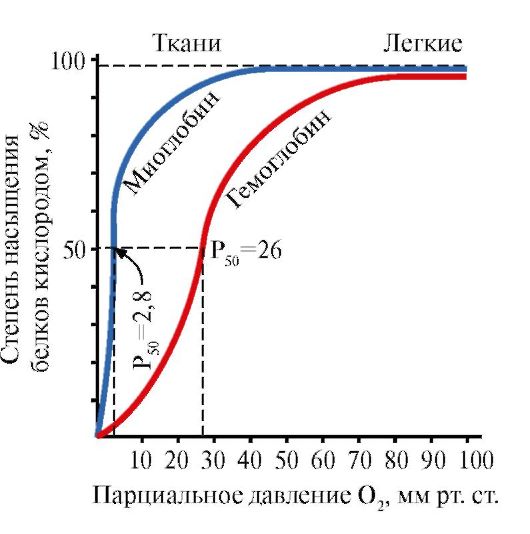 Rijst. 1.34. Verzadigingsafhankelijkheid van Мв enHbzuurstof uit zijn partiële druk
Rijst. 1.34. Verzadigingsafhankelijkheid van Мв enHbzuurstof uit zijn partiële druk
Merk op dat de vorm van de zuurstofverzadigingskrommen voor eiwitten anders is: voor myoglobine - hyperbool, voor hemoglobine - sigmoïde vorm.
1. Vergelijk de waarden van de partiële zuurstofdruk, waarbij MB en HB met 50% verzadigd zijn met O2. Welke van deze eiwitten heeft een hogere affiniteit voor O 2?
2. Welke structurele kenmerken van MB bepalen de hoge affiniteit voor О 2?
3. Welke structurele kenmerken van HB maken het mogelijk om O 2 af te geven in de haarvaten van rustende weefsels (bij een relatief hoge partiële druk van O 2) en deze terugkeer in werkende spieren sterk te verhogen? Welke eigenschap van oligomere eiwitten zorgt voor dit effect?
4. Bereken welke hoeveelheid O 2 (in%) zuurstofrijk hemoglobine geeft aan een rustende en werkende spier?
5. conclusies trekken over de relatie tussen de structuur van het eiwit en zijn functie.
2. De hoeveelheid zuurstof die door hemoglobine in haarvaten wordt afgegeven, hangt af van de intensiteit van katabole processen in weefsels (Bohr-effect). Hoe reguleren veranderingen in het weefselmetabolisme de affiniteit van Hb voor O 2? Effect van CO 2 en H + op de affiniteit van Hb voor O 2
 1. Beschrijf het Bohr-effect.
1. Beschrijf het Bohr-effect.
2. in welke richting is het proces weergegeven in het diagram:
a) in de haarvaten van de longen;
b) in weefselcapillairen?
3. Wat is de fysiologische betekenis van het Bohr-effect?
4. Waarom verandert de interactie van Hb met H+ op plaatsen ver van het heem de affiniteit van het eiwit voor O 2?
3. De affiniteit van Hb tot O2 hangt af van de concentratie van zijn ligand, 2,3-bisfosfoglyceraat, dat een allosterische regulator is van de affiniteit van Hb tot O2. Waarom beïnvloedt de interactie van het ligand op een plaats ver van de actieve plaats de functie van het eiwit? Hoe reguleert 2,3-BFG de affiniteit van Hb voor O 2? Beantwoord de volgende vragen om het probleem op te lossen:
1. Waar en waaruit wordt 2,3-bisfosfoglyceraat (2,3-BFG) gesynthetiseerd? Schrijf de formule op, geef de lading van een bepaald molecuul aan.
2. Met welke vorm van hemoglobine (oxy of deoxy) heeft BFH een wisselwerking en waarom? In welk deel van het HB-molecuul vindt de interactie plaats?
3. in welke richting is het proces weergegeven in het diagram
a) in weefselcapillairen;
b) in de haarvaten van de longen?
4.waar de concentratie van het complex hoger zou moeten zijn
NV-2,3-BFG:
a) in de haarvaten van spieren in rust,
b) in de haarvaten van werkende spieren (uitgaande van dezelfde concentratie BFG in erytrocyten)?
5. Hoe zal de affiniteit van Hb voor zuurstof veranderen wanneer een persoon zich aanpast aan omstandigheden op grote hoogte, als de concentratie van BFG in erytrocyten toeneemt? Wat is de fysiologische betekenis van dit fenomeen?
4. Vernietiging van 2,3-BFG tijdens opslag van ingeblikt bloed verstoort de functie van HB. Hoe zal de affiniteit van Hb tot O 2 in geconserveerd bloed veranderen als de concentratie van 2,3-BFG in erytrocyten kan dalen van 8 naar 0,5 mmol/l. Is het mogelijk om dergelijk bloed toe te dienen aan ernstig zieke patiënten als de concentratie van 2,3-BFG niet eerder dan drie dagen later is hersteld? Is het mogelijk om de functie van erytrocyten te herstellen door 2,3-BFG aan het bloed toe te voegen?
5. Denk aan de structuur van de eenvoudigste moleculen van immunoglobulinen. Welke rol spelen immunoglobulinen in het immuunsysteem? Waarom worden Ig vaak bivalenten genoemd? Hoe is de structuur van Ig gerelateerd aan hun functie? (Beschrijf een voorbeeld van een klasse van immunoglobulinen.)
Fysisch-chemische eigenschappen van eiwitten en methoden voor hun scheiding.
6. Hoe beïnvloedt de nettolading van een eiwit de oplosbaarheid ervan?
a) bepaal de totale lading van het peptide bij pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) hoe de lading van dit peptide zal veranderen bij pH> 7, pH<7, рН <<7?
c) wat is het iso-elektrisch punt van een eiwit (IEP) en in welke omgeving ligt het?
IEP van dit peptide?
d) bij welke pH-waarde zal de minste oplosbaarheid van dit peptide worden waargenomen.
7. Waarom "stremt" zure melk, in tegenstelling tot verse melk, tijdens het koken (dwz caseïnemelkeiwit slaat neer)? In verse melk hebben caseïnemoleculen een negatieve lading.
8. De gelfiltratiemethode wordt gebruikt om individuele eiwitten te scheiden. Een mengsel dat de eiwitten A, B, C met molecuulgewichten van respectievelijk 160.000, 80.000 en 60.000 bevat, werd geanalyseerd door gelfiltratie (Fig. 1.35). De gezwollen gelkorrels zijn doorlaatbaar voor eiwitten met een molecuulgewicht van minder dan 70.000. Wat is het principe achter deze scheidingsmethode? Welke grafiek geeft de fractioneringsresultaten correct weer? Specificeer de volgorde waarin eiwitten A, B en C uit de kolom komen.
 Rijst. 1.35. Een gelfiltratiemethode gebruiken om eiwitten te scheiden
Rijst. 1.35. Een gelfiltratiemethode gebruiken om eiwitten te scheiden
9. In afb. 1.36, A toont een diagram van elektroforese op papier van bloedserumeiwitten van een gezond persoon. De relatieve hoeveelheden eiwitfracties die met deze methode worden verkregen zijn: albuminen 54-58%, α 1-globulinen 6-7%, α 2-globulinen 8-9%, β-globulinen 13%, γ-globulinen 11-12%. ..
 Rijst. 1.36 Elektroforese op papier van bloedplasma-eiwitten van een gezond persoon (A) en een patiënt (B)
Rijst. 1.36 Elektroforese op papier van bloedplasma-eiwitten van een gezond persoon (A) en een patiënt (B)
I - -globulinen; II - -globulinen; III-α 2-globuline; IV-α 2-globuline; V - albumine
Veel ziekten gaan gepaard met kwantitatieve veranderingen in de samenstelling van serumeiwitten (dysproteïnemie). Bij het stellen van de diagnose en het beoordelen van de ernst en het stadium van de ziekte wordt rekening gehouden met de aard van deze veranderingen.
Gebruik de gegevens in de tabel. 1.5, maak een aanname over de ziekte, die wordt gekenmerkt door het elektroforetische profiel dat wordt weergegeven in Fig. 1.36.
Tabel 1.5. Veranderingen in de concentratie van serumeiwitten in pathologie
Eiwit actief centrum - het is de ligand-eiwitbindingsplaats. Op het oppervlak van de globule wordt een plaats gevormd die zich aan zichzelf kan hechten aan andere moleculen, de zogenaamde liganden ... Het actieve centrum van een eiwit wordt gevormd uit zijgroepen van aminozuren, dicht op het niveau van de tertiaire structuur. In de lineaire sequentie van de peptideketen kunnen ze zich op een significante afstand van elkaar bevinden. Eiwitten vertonen een hoge specificiteit bij interactie met een ligand. De hoge specificiteit van de interactie van het eiwit met het ligand wordt verzekerd door de complementariteit van de structuur van het actieve centrum van het eiwit met de structuur van het ligand. complementariteit Is de ruimtelijke en chemische correspondentie van op elkaar inwerkende moleculen. De ligand-eiwit bindingsplaatsen bevinden zich vaak tussen domeinen (de trypsine-ligand bindingsplaats heeft bijvoorbeeld 2 domeinen gescheiden door een groef).
De werking van eiwitten is gebaseerd op hun specifieke interactie met liganden. 50.000 individuele eiwitten die unieke actieve centra bevatten die alleen aan specifieke liganden kunnen binden en, vanwege de structurele kenmerken van het actieve centrum, hun inherente functies kunnen vertonen. Uiteraard bevat de primaire structuur informatie over de functie van eiwitten.
Quaternaire structuur- dit is het hoogste niveau van structurele organisatie, wat niet voor alle eiwitten mogelijk is. Onder een quaternaire structuur wordt verstaan een werkwijze voor het vouwen van polypeptideketens in de ruimte en de vorming van een enkele macromoleculaire formatie in structureel en functioneel opzicht. Elke afzonderlijk genomen polypeptideketen, genaamd protomeer of subeenheden , heeft meestal geen biologische activiteit. Het eiwit verwerft dit vermogen met een bepaalde methode van ruimtelijke associatie van zijn samenstellende protomeren. Het resulterende molecuul wordt meestal genoemd oligomeer (multimeer) .
De quaternaire structuur wordt gestabiliseerd door niet-covalente bindingen die ontstaan tussen de contactgebieden van protomeren, die op complementaire wijze met elkaar interageren.
Quaternaire eiwitten omvatten veel enzymen (lactaatdehydrogenase, glutamaatdehydrogenase, enz.), evenals hemoglobine, het spiercontractiele eiwit myosine. Sommige eiwitten hebben een klein aantal subeenheden 2 - 8, andere honderden en zelfs duizenden subeenheden. Het eiwit van het tabaksmozaïekvirus heeft bijvoorbeeld 2.130 subeenheden.
Een typisch voorbeeld van een eiwit met een quaternaire structuur is hemoglobine. Het hemoglobinemolecuul bestaat uit 4 subeenheden, dat wil zeggen polypeptideketens, die elk zijn geassocieerd met heem, waarvan 2 polypeptideketens -2afla en -2beta worden genoemd. Ze verschillen in de primaire structuur en de lengte van de polypeptideketen.
De bindingen die de quaternaire structuur vormen zijn minder sterk. Onder invloed van sommige middelen wordt het eiwit verdeeld in afzonderlijke subeenheden. Wanneer het middel wordt verwijderd, kunnen de subeenheden zich herenigen en wordt de biologische functie van het eiwit hersteld. Dus wanneer ureum wordt toegevoegd aan de hemoglobine-oplossing, valt het uiteen in 4 subeenheden, en wanneer ureum wordt verwijderd, wordt de structurele en functionele rol van hemoglobine hersteld.
Einde van het werk -
Dit onderwerp hoort bij de sectie:
Biochemie. Eiwitten. Aminozuren - structurele componenten van eiwitten
Eiwitten aminozuren zijn de structurele componenten van eiwitten .. eiwitten .. eiwitten zijn stikstofhoudende hoogmoleculaire organische verbindingen bestaande uit aminozuren die in een keten met ..
Als u aanvullend materiaal over dit onderwerp nodig heeft, of u heeft niet gevonden wat u zocht, raden we u aan de zoekopdracht in onze database te gebruiken:
Wat doen we met het ontvangen materiaal:
Als dit materiaal nuttig voor u bleek te zijn, kunt u het opslaan op uw pagina op sociale netwerken:
| Tweeten |
Alle onderwerpen in deze sectie:
Enzym werkingsmechanisme
Volgens moderne concepten kunnen tijdens de interactie van een enzym met een substraat conventioneel 3 stadia worden onderscheiden: Stadium 1 wordt gekenmerkt door diffusie van het substraat naar het enzym.
Zuur-base katalyse
Het actieve centrum van het enzym bevat zure en basische groepen. Zuurgroepen splitsen H+ en hebben een negatieve lading. Groepen van het basistype voegen H + toe en hebben een polo
EEN). Fisher's hypothese
Volgens het is er een strikte sterische overeenkomst tussen het substraat en het actieve centrum van het enzym. Volgens Fischer is een enzym een starre structuur en is een substraat als het ware een afgietsel van zijn actieve cent
Koolhydraat metabolisme
UITWISSELING VAN KOOLHYDRATEN 1. De belangrijkste koolhydraten van het dierlijke organisme, hun biologische rol. 2. Omzetting van koolhydraten in de organen van het spijsverteringsstelsel. 3. Biosynthese en verval
De biologische rol van koolhydraten
BIOLOGISCHE ROL VAN KOOLHYDRATEN: 1. ENERGIE. Wanneer 1 g koolhydraten wordt geoxideerd tot eindproducten (CO2 en H2O), komt 4,1-kcal energie vrij. Het aandeel koolhydraten is ongeveer 60-70
Omzetting van koolhydraten in het spijsverteringskanaal
OMZETTING VAN KOOLHYDRATEN IN HET SPIJSVERTERINGSKANAAL De belangrijkste koolhydraten van voedsel voor het menselijk lichaam zijn: zetmeel, glycogeen, sucrose, lactose. ingenomen zetmeel
Biosynthese en afbraak van glycogeen
BIOSYNTHESE EN AFBRAAK VAN GLYCOGEN IN WEEFSELS. GLYCOGEN ZIEKTEN. Het bleek dat glycogeen in bijna alle organen en weefsels kan worden gesynthetiseerd. Echter, het grootste einde
Anaërobe glycolyse
Afhankelijk van de functionele toestand van het lichaam, kunnen de cellen van organen en weefsels ofwel in omstandigheden met voldoende zuurstoftoevoer zijn of het gebrek ervan ervaren, dan
Aerobe glycolyse (hexosedifosfaatroute)
HEXOSODIFOSFAAT MANIER. Deze klassieke route van aëroob katabolisme van koolhydraten in weefsels verloopt in het cytoplasma tot aan het stadium van pyruvaatvorming en eindigt in mitochondriën met de vorming van con
Hexose monofosfaat route
HEXOSOMONOFOSFAAT PAD VAN GLUCOSE CONVERSIE IN WEEFSELS, CHEMIE VAN REACTIES. Oxidatie van glucose langs deze route vindt plaats in het cytoplasma van cellen en wordt weergegeven door twee opeenvolgende takken
Gluconeogenese
GLUCONEOGENESE De belangrijkste bronnen van glucose voor het menselijk lichaam zijn: 1. koolhydraten in de voeding; 2. weefselglycogeen; 3. gluconeogenese. GLUCONEOGENESE is
De belangrijkste lipiden van het menselijk lichaam en hun biologische rol
LIPIDEN worden complexe organische stoffen van biologische aard genoemd, onoplosbaar in water, maar oplosbaar in organische oplosmiddelen. LIPIDEN zijn het hoofdvoedsel. ze zijn nee
Lipidenvertering, vethersynthese
Vertering van lipiden. LIPIDEN die met voedsel in de mondholte worden geleverd, ondergaan alleen mechanische verwerking. LIPOLYTISCHE enzymen worden niet gevormd in de mondholte. Vertering van vet
Bloedlipoproteïnen
LIPIDEN zijn in water onoplosbare verbindingen, dus voor hun bloedtransport zijn speciale transporters nodig die in water oplosbaar zijn. Deze transportvormen zijn LIPO-EIWITTEN.
Oxidatie van hogere vetzuren
Vetweefsel, bestaande uit adipocyten, speelt een specifieke rol in de vetstofwisseling. Ongeveer 65% van de massa vetweefsel valt op het aandeel triacylglycerolen (TAG) dat erin is afgezet - ze vertegenwoordigen
Biosynthese van IVH in weefsels
De biosynthese van IVA vindt plaats in het endoplasmatisch reticulum van cellen. Vervangbare IVA (allemaal beperkend en onverzadigd, met één dubbele binding) worden gesynthetiseerd in cellen uit ACETYL-CoA. Voorwaarden voor bi
cholesterol uitwisseling
Cholesterol uitwisseling. Cholesterol is een voorloper in de synthese van steroïden: galzuren, steroïde hormonen, vitamine D 3. Cholesterol is een essentieel structureel bestanddeel
Vertering van eiwitten
Vertering van eiwitten in het spijsverteringskanaal Voedingseiwitten ondergaan hydrolytische afbraak onder invloed van PROTEOLYTISCHE ENZYMEN (klasse - hydrolasen, subklasse - peptidasen).
Verval van aminozuren, neutralisatie van vervalproducten
ROTATIE VAN AMINOZUREN Aminozuren die niet zijn opgenomen komen in de dikke darm terecht, waar ze gaan rotten. ROTATIE VAN AMINOZUREN is het proces van afbraak van aminozuren onder invloed van:
Aminozuurmetabolisme
Aminozuurmetabolisme De bronnen van aminozuren in de cel zijn: 1. voedingseiwitten na hun hydrolyse in de spijsverteringsorganen; 2. synthese van niet-essentiële aminozuren;
Manieren om ammoniak te neutraliseren
Ammoniak wordt gevormd uit aminozuren tijdens de afbraak van andere stikstofhoudende verbindingen (biogene aminen, NUCLEOTIDEN). Veel van de ammoniak wordt gevormd in de dikke darm tijdens verrotting. Het wordt opgenomen in de cr
Regulering van de stofwisseling
SIGNAALMOLECULEN. De belangrijkste taken van de regulering van het metabolisme en cellulaire functies: 1. intracellulaire en intercellulaire coördinatie van metabolische processen; 2. de uitsluiting van "inactief"
Hypothalamische hormonen
HYPOTHALAMUS HORMONEN De HYPOTHALAMUS is een component en een soort "uitlaat" van het limbische systeem. Dit is een deel van het diencephalon dat verschillende parameters van gom . regelt
hypofyse hormonen
hypofyse hormonen
Hypofysehormonen De anterieure (adenohypofyse) en achterste lobben (neurohypofyse) worden uitgescheiden in de hypofyse. De hormonen van de adenohypofyse kunnen worden onderverdeeld in 3 groepen, afhankelijk van:
Biosynthese van joodthyronines
De synthese van joodthyronines vindt plaats in de samenstelling van het eiwit - thyroglobuline, dat zich in de follikels van de schildklier bevindt. Thyroglobuline is een glycoproteïne dat 115 tyrosineresiduen bevat. P
lipidenmetabolisme
In de lever stimuleren vetweefselhormonen de lipolyse. Deze effecten op het metabolisme van koolhydraten en lipiden gaan gepaard met een toename van de gevoeligheid van cellen voor de werking van adrenaline onder invloed van schildklierhormonen.
Hyposecretie
In de kindertijd leidt een afname van de secretie tot een vertraging in de fysieke en mentale ontwikkeling (cretinisme). Bij volwassenen is een ernstige manifestatie van een tekort aan schildklierhormonen een mix
hypersecretie
Diffuus toxisch struma (ziekte van Graves) is de meest voorkomende ziekte, die gepaard gaat met een verhoogde productie van joodthyronines. Bij deze ziekte wordt de grootte van de schildklier vergroot en
bijschildklierhormonen
Parathyroïdhormoon wordt gesynthetiseerd in de bijschildklieren en bestaat uit 84 aminozuurresiduen. Het hormoon wordt opgeslagen in secretoire korrels. De secretie van PTH wordt gereguleerd door het calciumgehalte in het bloed:
geslachtshormonen
Gonadale hormonen Door hun chemische aard zijn het steroïden. Toewijzen: 1. Androgenen; 2. Oestrogenen; 3. Progestagenen.
Bijnier hormonen
Bijnierhormonen De bijnieren zijn endocriene klieren die de cortex en medulla afscheiden. In de corticale laag worden steroïde hormonen gesynthetiseerd, in de hersenen
Pancreashormonen
Pancreashormonen Functies van de alvleesklier: · exocrien; endocrien. De exocriene functie bestaat uit de synthese en afscheiding van spijsverteringsfer
examenvragen
FARMACEUTISCHE FACULTEIT (CORRESPONDENTIEAFDELING) Examenvragen in de biologische chemie voor studenten van het 3e jaar (6e semester) 1. Biochemie, haar taken. De relatie van biochemie met ph
EIWITMOLECULE CONFIGURATIE EN CONFORMATIE
Elektronenmicroscopie
Het kan worden gebruikt om de structuur van eiwitmoleculen met een groot molecuulgewicht te verduidelijken - van 500.000 tot 1.000.000 Da (dalton). Dalton (Ja) en kilodalton (kDa)- meeteenheden van de massa van eiwitten. 1 kDa = 10 3 Ja. 1 dalton is gelijk aan 1/16 van de massa van een zuurstofatoom (zuurstofmassa-eenheid).
Uit alles wat gezegd is, kunnen we concluderen dat de ruimtelijke organisatie van eiwitten erg complex is. In de chemie is er een concept - ruimtelijk CONFIGURATIE - star gefixeerd door covalente bindingen, de ruimtelijke onderlinge rangschikking van de delen van het molecuul(bijvoorbeeld: behorend tot de L-serie van stereo-isomeren of tot de D-serie).
Voor eiwitten wordt het concept ook gebruikt CONFORMATIE een eiwitmolecuul - een welomlijnde, maar niet bevroren, niet constante relatieve positie van de delen van het molecuul... Omdat de conformatie van een eiwitmolecuul wordt gevormd met de deelname van zwakke soorten bindingen, is het mobiel (in staat tot veranderingen) en kan het eiwit zijn structuur veranderen. Afhankelijk van de omstandigheden van de externe omgeving, kan het molecuul in verschillende conformatietoestanden bestaan, die gemakkelijk in elkaar overgaan. Slechts één of meerdere conformatietoestanden, waartussen een evenwicht bestaat, zijn energetisch gunstig voor reële omstandigheden. Overgangen van de ene conformationele toestand naar de andere zorgen voor de werking van het eiwitmolecuul. Dit zijn omkeerbare conformatieveranderingen (ze treden bijvoorbeeld op in het lichaam tijdens de geleiding van een zenuwimpuls, tijdens de overdracht van zuurstof door hemoglobine). Wanneer de conformatie verandert, worden enkele van de zwakke bindingen vernietigd en worden nieuwe zwakke bindingen gevormd.
De interactie van een eiwit met een bepaalde stof leidt soms tot de binding van een molecuul van deze stof aan een eiwitmolecuul. Dit fenomeen staat bekend als "Sorptie" (bindend)... Het omgekeerde proces - de afgifte van een ander molecuul uit het eiwit dat men heet "Desorptie".
Als voor een paar moleculen het sorptieproces prevaleert boven desorptie, dan is dit al: specifieke sorptie, en de stof die wordt gesorbeerd heet "ligand".
Soorten liganden:
1) Eiwit-enzym ligand - substraat.
2) Transporteiwitligand - getransporteerde stof.
3) Antilichaam (immunoglobuline) ligand - antigeen.
4) Hormoonreceptor of neurotransmitterligand - hormoon of neurotransmitter.
Een eiwit kan zijn conformatie veranderen, niet alleen bij interactie met een ligand, maar ook als gevolg van elke chemische interactie. Een voorbeeld van een dergelijke interactie is de toevoeging van een fosforzuurresidu.
Onder natuurlijke omstandigheden hebben eiwitten verschillende thermodynamisch gunstige conformatietoestanden. Dit zijn inheemse staten (natuurlijk). Natura (lat.) - natuur.


