Trung tâm hoạt động của protein và sự tương tác của nó với phối tử. Chức năng của protein
Trung tâm hoạt động của protein là một vùng nhất định của phân tử protein, thường nằm ở phần lõm của nó, được hình thành bởi các gốc axit amin được tập hợp trong một vùng không gian nhất định trong quá trình hình thành cấu trúc bậc ba và có khả năng liên kết bổ sung với phối tử. Trong trình tự tuyến tính của chuỗi polypeptide, các gốc tạo thành trung tâm hoạt động có thể nằm ở một khoảng cách đáng kể với nhau.
Độ đặc hiệu cao của liên kết protein với phối tử được đảm bảo bởi tính bổ sung giữa cấu trúc của trung tâm hoạt động của protein và cấu trúc của phối tử.
Tính bổ sung đề cập đến sự tương ứng về không gian và hóa học của các phân tử tương tác. Phối tử phải có khả năng đi vào và trùng khớp về mặt không gian với cấu trúc của vị trí hoạt động. Sự trùng khớp này có thể không hoàn chỉnh, nhưng do tính không ổn định về hình dạng của protein, vị trí hoạt động có thể có những thay đổi nhỏ và được “điều chỉnh” theo phối tử. Ngoài ra, giữa các nhóm chức của phối tử và các gốc axit amin tạo thành trung tâm hoạt động phải phát sinh các liên kết giữ phối tử ở trung tâm hoạt động. Liên kết giữa phối tử và trung tâm hoạt động của protein có thể là không cộng hóa trị (ion, hydro, kỵ nước) hoặc cộng hóa trị.
Đặc điểm của trung tâm hoạt động
Trung tâm hoạt động của protein là vùng tương đối cách ly với môi trường xung quanh protein, được hình thành bởi các gốc axit amin. Trong khu vực này, mỗi phần cặn, do quy mô riêng và các nhóm chức năng của nó, tạo thành “khung đỡ” của trung tâm hoạt động.
Tính chất độc đáo trung tâm hoạt động không chỉ phụ thuộc vào tính chất hóa học của các axit amin tạo thành nó mà còn phụ thuộc vào sự định hướng tương đối chính xác của chúng trong không gian. Do đó, ngay cả những vi phạm nhỏ về cấu trúc chung của protein do thay đổi điểm trong cấu trúc hoặc điều kiện bậc một của nó môi trường có thể dẫn đến những thay đổi về tính chất hóa học và chức năng của các gốc hình thành nên trung tâm hoạt động, phá vỡ sự liên kết của protein với phối tử và chức năng của nó. Trong quá trình biến tính, trung tâm hoạt động của protein bị phá hủy và hoạt động sinh học của chúng bị mất.
Thông thường, trung tâm hoạt động được hình thành theo cách mà khả năng tiếp cận của nước với các nhóm chức năng của các gốc của nó bị hạn chế, tức là. điều kiện được tạo ra để liên kết phối tử với các gốc axit amin.
Vị trí liên kết phối tử protein thường nằm giữa các miền. Ví dụ, enzyme phân giải protein trypsin, tham gia vào quá trình thủy phân liên kết peptide của protein thực phẩm trong ruột, có 2 miền được phân tách bằng một rãnh. Bề mặt bên trong của rãnh được hình thành bởi các gốc axit amin của các miền này, nằm cách xa nhau trong chuỗi polypeptide (Ser 177, His 40, Asp 85).
Các vùng khác nhau trong protein có thể di chuyển tương đối với nhau khi tương tác với phối tử, điều này tạo điều kiện thuận lợi cho protein tiếp tục hoạt động. Đặc tính chính của protein làm cơ sở cho chức năng của chúng là tính chọn lọc trong việc gắn các phối tử cụ thể vào các phần nhất định của phân tử protein.
Các loại phối tử:
Các phối tử có thể là chất vô cơ (thường là ion kim loại) và chất hữu cơ, chất có trọng lượng phân tử thấp và chất có trọng lượng phân tử cao;
Có những phối tử thay đổi cấu trúc hóa học khi gắn vào vị trí hoạt động của protein (làm thay đổi cơ chất ở vị trí hoạt động của enzyme);
Có những phối tử chỉ gắn vào protein tại thời điểm hoạt động (ví dụ, O 2 được vận chuyển bởi huyết sắc tố) và các phối tử liên tục liên kết với protein và đóng vai trò phụ trợ trong hoạt động của protein (ví dụ: sắt, là một phần của huyết sắc tố).
CHƯƠNG 3
ENZYM. CƠ CHẾ HOẠT ĐỘNG CỦA ENZYME
Enzyme hoặc enzyme là các protein cụ thể là một phần của tất cả các tế bào và mô của sinh vật sống và hoạt động như chất xúc tác sinh học.
Tính chất chung của enzyme và chất xúc tác vô cơ:
1. Chúng không bị tiêu hao trong quá trình phản ứng.
2. Chúng phát huy tác dụng ở nồng độ thấp.
3. Chúng không ảnh hưởng đến giá trị hằng số cân bằng của phản ứng.
4. Hành động của họ tuân theo quy luật hành động quần chúng.
5. Không tăng tốc các phản ứng không thể thực hiện được về mặt nhiệt động.
Sự khác biệt giữa enzyme và chất xúc tác vô cơ.
1. Tính bền nhiệt của enzyme.
2. Sự phụ thuộc hoạt động của enzyme vào pH môi trường.
3. Tính đặc hiệu của hoạt động của enzyme.
4. Tốc độ phản ứng enzym tuân theo các định luật động học nhất định.
5. Hoạt động của enzyme phụ thuộc vào hoạt động của chất điều hòa - chất kích hoạt và chất ức chế.
6. Một số enzyme trải qua quá trình biến đổi sau tổng hợp trong quá trình hình thành cấu trúc bậc ba và bậc bốn.
7. Kích thước của các phân tử enzyme thường lớn hơn nhiều so với kích thước cơ chất của chúng.
Cấu trúc phân tử enzyme
Theo cấu trúc, enzyme có thể là protein đơn giản hoặc phức tạp. Enzym là một protein phức tạp được gọi là holoenzim. Phần protein của enzyme được gọi là apoenzym, phần không phải protein được gọi là cofactor. Phân biệt hai loại đồng yếu tố:
1. Nhóm giả - liên kết chặt chẽ với apoenzym, thường bằng liên kết cộng hóa trị.
2. Coenzym là phần không phải protein, dễ dàng tách ra khỏi apoenzim. Các dẫn xuất của vitamin thường được sử dụng làm coenzym.
Đến coenzym Các kết nối sau bao gồm:
Dẫn xuất vitamin;
Hemes, là một phần của cytochromes, catalase, peroxidase, guanylate cyclase, NO synthase và là một nhóm enzyme giả;
Nucleotide là chất cho và chất nhận dư lượng axit photphoric;
Ubiquinone hoặc coenzym Q, tham gia vào quá trình chuyển electron và proton trong chuỗi hô hấp của mô;
Phosphoadenosylphosphosulfate, tham gia vận chuyển sunfat;
Glutathione, tham gia vào phản ứng oxi hóa khử.
Bảng 3.1.
Chức năng coenzym của vitamin
Ppt%5C34928-slozhnye_belki_ch1_1.jpg" alt=">Trung tâm hoạt động của protein và sự tương tác của nó với phối tử. Trong quá trình hình thành cấu trúc bậc ba"> Активный центр белка и его взаимодействие с лигандом. В процессе формирования третичной структуры на поверхности функционально активного белка, обычно в углублении, образуется участок, сформированный радикалами аминокислот, далеко стоящими друг от друга в первичной структуре. Этот участок, имеющий уникальное строение для данного белка и способный специфично взаимодействовать с определенной молекулой или группой похожих молекул, называется центром связывания белка с лигандом или активным центром. Лигандами называются молекулы, взаимодействующие с белками.!}
Ppt%5C34928-slozhnye_belki_ch1_2.jpg" alt=">Một phối tử có thể là chất có trọng lượng phân tử thấp hoặc chất có trọng lượng phân tử cao (đại phân tử), bao gồm"> Лигандом может быть как низкомолекулярное, так и высокомолекулярное (макромолекула) вещество, в том числе и другой белок. Лигандами являются субстраты ферментов, кофакторы, ингибиторы и активаторы ферментов, протомеры в олигомерном белке и т.д.!}
Ppt%5C34928-slozhnye_belki_ch1_3.jpg" alt=">Tính đặc hiệu cao của tương tác phối tử-protein được đảm bảo bởi tính bổ sung giữa cấu trúc của trung tâm hoạt động với cấu trúc của phối tử.">!}
Ppt%5C34928-slozhnye_belki_ch1_4.jpg" alt=">Tính bổ sung là sự tương ứng về không gian và hóa học của các bề mặt tương tác. Trung tâm hoạt động không chỉ"> Комплементарность - это пространственное и химическое соответствие взаимодействующих поверхностей. Активный центр должен не только пространственно соответствовать входящему в него лиганду, но и между функциональными группами радикалов, входящих в активный центр, и лигандом должны образоваться связи чаще всего нековалентные (ионные, водородные, а также гидрофобные взаимодействия), которые удерживают лиганд в активном центре.!}
Ppt%5C34928-slozhnye_belki_ch1_5.jpg" alt=">Tương tác bổ sung của protein với phối tử">!}
Ppt%5C34928-slozhnye_belki_ch1_6.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_7.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_8.jpg" alt="> PHÂN LOẠI PROTEIN 1. Protein đơn giản chỉ bao gồm các axit amin. 2. Protein phức tạp (holoprotein)"> КЛАССИФИКАЦИЯ БЕЛКОВ 1. Простые белки состоят только из аминокислот. 2. Сложные белки (холопротеины) содержат белковую часть (апопротеин) и небелковую (простетическую) группу.!}
Ppt%5C34928-slozhnye_belki_ch1_9.jpg" alt=">Các chất hữu cơ (lipid, carbohydrate) và vô cơ (kim loại) khác nhau có thể hoạt động như một nhóm chân tay giả."> В качестве простетической группы могут выступать различные органические (липиды, углеводы) и неорганические (металлы) вещества. Связь между простетической группой и апопротеином может быть как ковалентная, так и нековалентная. Простетическую группу порой можно рассматривать в качестве лиганда. Наличие небелковой части обеспечивает выполнение белком его функции. При утрате простетической группы холопротеин теряет свою активность.!}
Ppt%5C34928-slozhnye_belki_ch1_10.jpg" alt=">Protein phức tạp - chromoprotein - nucleoprotein - lipoprotein - phosphoprotein - glycoprotein - metallicoprotein">!}
Ppt%5C34928-slozhnye_belki_ch1_11.jpg" alt=">Metalloprotein bao gồm các holoenzym chứa các ion kim loại phối hợp không phải heme. Trong số các metallicoprotein có protein"> Металлопротеинам можно отнести холоферменты, содержащие негемовые координационно связанные ионы металлов. Среди металлопротеинов есть белки, выполняющие депонирующие и транспортные функции (например, железосодержащие ферритин и трансферрин) и ферменты (например, цинксодержащая карбоангидраза и различные супероксиддисмутазы, содержащие в качестве активных центров ионы меди, марганца, железа и других металлов). Но и хромопротеины, содержащие ионы металлов, также можно отнести к металлопротеинам.!}
Ppt%5C34928-slozhnye_belki_ch1_12.jpg" alt=">Metalloprotein thường là enzyme. Các ion kim loại trong trường hợp này: - tham gia vào quá trình định hướng của chất nền"> Металлопротеины часто являются ферментами. Ионы металлов в этом случае: - участвуют в ориентации субстрата в активном центре фермента, входят в состав активного центра фермента и участвуют в катализе, являясь, например, акцепторами электронов на определенной стадии ферментативной реакции. Часто ион металла в составе фермента называют кофактором.!}
Ppt%5C34928-slozhnye_belki_ch1_13.jpg" alt=">Enzymatic metallicoprotein bao gồm các protein có chứa, ví dụ: - đồng - cytochrome oxidase, trong phức hợp"> К ферментативным металлопротеинам относятся белки, содержащие например: - медь – цитохромоксидаза, в комплексе с другими ферментами дыхательной цепи митохондрий участвует в синтезе АТФ, - железо – ферритин, депонирующий железо в клетке, трансферрин, переносящий железо в крови, каталаза, обезвреживающая перекись водорода, - цинк – алкогольдегидрогеназа, обеспечивающая метаболизм этанола и других спиртов, лактатдегидрогеназа, участвующая в метаболизме молочной кислоты, - карбоангидраза, образующая угольную кислоту из CO2 и H2O, - щелочная фосфатаза, гидролизующая фосфорные эфиры !} kết nối khác nhau, - α2-macroglobulin, protein kháng protease trong máu. - selen - peroxidase tuyến giáp, tham gia tổng hợp hormone tuyến giáp, enzyme chống oxy hóa glutathione peroxidase, - canxi - α-amylase của nước bọt và dịch tụy, thủy phân tinh bột.
Ppt%5C34928-slozhnye_belki_ch1_14.jpg" alt=">Ferritin">!}
Ppt%5C34928-slozhnye_belki_ch1_15.jpg" alt=">Phosphoprotein là các protein có chứa nhóm phốt phát. Nó liên kết với chuỗi peptide"> Фосфопротеины – это белки, в которых присутствует фосфатная группа. Она связывается с пептидной цепью через остатки тирозина, серина и треонина, т.е. тех аминокислот, которые содержат ОН-группу. Способ присоединения фосфата к белку на примере серина и тирозина!}
Ppt%5C34928-slozhnye_belki_ch1_16.jpg" alt=">Axit photphoric có thể thực hiện: - Vai trò cấu trúc, truyền điện tích, độ hòa tan và thay đổi tính chất"> Фосфорная кислота может выполнять: - Структурную роль, придавая заряд, растворимость и изменяя свойства белка, например, в казеине молока, яичном альбумине. Наличие остатков фосфорной кислоты способствует связыванию кальция, что необходимо для формирования, например, костной ткани. - Функциональную роль. В клетке присутствует много белков, которые связаны с фосфатом не постоянно, а в зависимости от активности метаболизма. Белок может многократно переходить в фосфорилированную или в дефосфорилированную форму, что играет регулирующую роль в его работе.!}
Ppt%5C34928-slozhnye_belki_ch1_17.jpg" alt=">Phosphoryl hóa là quá trình chuyển dư lượng axit photphoric từ chất cho phosphoryl hóa sang chất nền, thường là"> Фосфорилирование - процесс переноса остатка фосфорной кислоты от фосфорилирующего агента-донора к субстрату, как правило, катализируемый ферментами (киназами) и ведущий к образованию эфиров фосфорной кислоты. Дефосфорилирование (утрату остатка фосфорной кислоты) катализируют фосфатазы. АТФ + R-OH → АДФ + R-OPO3H2 R-OPO3H2 + Н2О → R-OH + Н3РО4!}
Ppt%5C34928-slozhnye_belki_ch1_18.jpg" alt=">Ví dụ: 1) enzyme glycogen synthase và glycogen phosphorylase 2) histone ở trạng thái phosphoryl hóa liên kết kém chặt chẽ hơn"> Примеры: 1) ферменты гликогенсинтаза и гликогенфосфорилаза 2) гистоны в фосфорилированном состоянии менее прочно связываются с ДНК и активность генома возрастает. Изменение конформации белка в фосфорилированном и дефосфорилированном состоянии!}
Ppt%5C34928-slozhnye_belki_ch1_19.jpg" alt=">Lipoprotein chứa lipid không liên kết cộng hóa trị như một bộ phận giả. Đặc biệt là lipid"> Липопротеины содержат в качестве простетической части нековалентно связанные липиды. Липиды, в частности жиры, холестерол и его эфиры не растворяются в водных фазах организма, поэтому транспорт их кровью и лимфой осуществляется в виде комплексов с белками и фосфолипидами, которые называются липопротеинами.!}
Ppt%5C34928-slozhnye_belki_ch1_20.jpg" alt=">Tất cả các lipoprotein đều có cấu trúc tương tự nhau: lõi bao gồm các phân tử kỵ nước: triacylglycerol, este cholesterol và"> Все липопротеины имеют сходное строение: ядро состоит из гидрофобных молекул: триацилглицеролов, эфиров холестерола, а на поверхности находится монослой фосфолипидов, полярные группы которых обращены к воде, а гидрофобные погружены в гидрофобное ядро липопротеина. Кроме фосфолипидов, на поверхности находятся белки – аполипопротеины (апобелками). Их выделяют несколько видов: А, В, С, D. В каждом типе липопротеинов преобладают соответствующие ему апобелки. Аполипопротеины выполняют различные функции. Интегральные аполипопротеины являются структурными компонентами. Периферические аполипопротеины в плазме крови могут передаваться от одного типа липопротеинов к другим, определяя их дальнейшие превращения.!}
Ppt%5C34928-slozhnye_belki_ch1_21.jpg" alt=">Sơ đồ cấu trúc của lipoprotein Cấu trúc của lipoprotein">!}
Ppt%5C34928-slozhnye_belki_ch1_22.jpg" alt=">Cấu trúc của lipoprotein huyết tương máu">!}
Ppt%5C34928-slozhnye_belki_ch1_23.jpg" alt=">Có bốn loại lipoprotein chính: - lipoprotein mật độ cao (HDL), - lipoprotein mật độ thấp (LDL),"> Выделяют четыре основных класса липопротеинов: -липопротеины высокой плотности (ЛПВП), -липопротеины низкой плотности (ЛПНП), -липопротеины очень низкой плотности (ЛПОНП), -хиломикроны (ХМ). Каждый из типов ЛП образуется в !} các loại vải khác nhau và vận chuyển một số lipid. Nồng độ và tỷ lệ của một số lipoprotein nhất định trong máu đóng vai trò hàng đầu trong việc xuất hiện một bệnh lý mạch máu phổ biến như xơ vữa động mạch. HDL có tác dụng chống xơ vữa động mạch, LDL và VLDL là chất gây xơ vữa động mạch.
Ppt%5C34928-slozhnye_belki_ch1_24.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_25.jpg" alt=">Glycoprotein hoặc glycoconjugates là các protein có chứa thành phần carbohydrate liên kết cộng hóa trị với xương sống polypeptide."> Гликопротеины или, гликоконъюгаты – это белки, содержащие углеводный компонент, ковалентно присоединенный к полипептидной основе. Содержание углеводов в них варьирует от 1% до 98% по массе. Два подкласса белков, содержащих углеводы: ■ протеогликаны ■ гликопротеины!}
Mô tả="">
Ppt%5C34928-slozhnye_belki_ch1_27.jpg" alt=">Glycoprotein được đặc trưng bởi hàm lượng carbohydrate thấp, được gắn: - bằng liên kết N-glycosid với nhóm NH2 của một số"> Для гликопротеинов характерно невысокое содержание углеводов, которые присоединены: - N-гликозидной связью к NН2-группе какого-нибудь аминокислотного остатка, например, аспарагина; - О-гликозидной связью к гидроксильной группе остатка серина, треонина,тирозина, гидроксилизина.!}
Ppt%5C34928-slozhnye_belki_ch1_28.jpg" alt=">Sự hình thành liên kết O- và N-glycosid trong glycoprotein. 1 - Liên kết N-glycosid giữa nhóm amide"> Образование О- и N-гликозидных связей в гликопротеинах. 1 - N-гликозидная связь между амидной группой аспарагина и ОН-группой моносахарида; 2 - О-гликозидная связь между ОН-группой серина и ОН-группой моносахарида.!}
Ppt%5C34928-slozhnye_belki_ch1_29.jpg" alt=">Phương pháp thêm carbohydrate vào protein">!}
Ppt%5C34928-slozhnye_belki_ch1_30.jpg" alt=">Chức năng của glycoprotein là: 1. Cấu trúc - thành tế bào vi khuẩn, ma trận xương, ví dụ như collagen, đàn hồi."> Функцией гликопротеинов являются: 1. Структурная – клеточная стенка бактерий, костный матрикс, например, коллаген, эластин. 2. Защитная – например, антитела, интерферон, факторы свертывания крови (протромбин, фибриноген). 3. Рецепторная – присоединение эффектора приводит к изменению конформации белка-рецептора, что вызывает внутриклеточный ответ. 4. Гормональная – гонадотропный, адренокортикотропный и тиреотропный гормоны. 5. Ферментативная – холинэстераза, нуклеаза. 6. Транспортная – перенос веществ в крови и через мембраны, например, трансферрин, транскортин, альбумин, Na+,К+-АТФаза.!}
Ppt%5C34928-slozhnye_belki_ch1_31.jpg" alt=">Sơ đồ cấu trúc của protein thụ thể">!}
Ppt%5C34928-slozhnye_belki_ch1_32.jpg" alt=">Chromoprotein là tên gọi chung của các protein phức tạp với các nhóm chân tay giả có màu có tính chất hóa học khác nhau."> Хромопротеины - собирательное название сложных белков с окрашенными простетическими группами различной химической природы. гемопротеины (содержат гем), ретинальпротеины (содержат витамин А), флавопротеины (содержат витамин В2), кобамидпротеины (содержат витамин В12).!}
Ppt%5C34928-slozhnye_belki_ch1_33.jpg" alt=">Flavoprotein là enzyme của phản ứng oxi hóa khử. Chúng chứa các dẫn xuất vitamin B2 flavin mononucleotide (FMN) và flavin adenine dinucleotide"> Флавопротеины - это ферменты окислительно-восстановительных реакций. Содержат производные витамина В2 флавинмононуклеотид (ФМН) и флавинадениндинуклеотид (ФАД). Связываются данные простетические группы ковалентно и придают желтое окрашивание. Эти простетические группы являются производными изоаллоксазина.!}
Ppt%5C34928-slozhnye_belki_ch1_34.jpg" alt=">Isoalloxazine là một hợp chất dị vòng, một dẫn xuất của pteridine. Phân tử isoalloxazine bao gồm ba vòng thơm -"> Изоаллоксазин - гетероциклическое соединения, производное птеридина. Молекула изоаллоксазина состоит из трех ароматических колец - бензольного, пиримидинового, пиразинового.!}
Ppt%5C34928-slozhnye_belki_ch1_35.jpg" alt=">Hemoprotein là các chromoprotein chứa heme. Là một thành phần phi protein, chúng bao gồm các porphyrin sắt hoặc magie có cấu trúc tương tự nhau."> Гемопротеины - гем-содержащие хромопротеины. В качестве небелкового компонента включают структурно сходные железо- или магнийпорфирины. Белковый компонент может быть разнообразным как по составу, так и по структуре. Основу структуры простетической группы большинства гемосодержащих белков составляет порфириновое кольцо, являющееся в свою очередь производным тетрапиррольного соединения – порфирина. Порфирин!}
Ppt%5C34928-slozhnye_belki_ch1_36.jpg" alt=">Vòng porphyrin có khả năng hình thành các hợp chất phối hợp với nhiều ion kim loại khác nhau. Là kết quả của sự tạo phức,"> Порфириновое кольцо способно образовывать координационные соединения с различными ионами металлов. В результате комплексообразования формируются металлопорфирины: содержащие ионы железа – гемоглобины, миоглобин, цитохромы, пероксидаза, каталаза и др. (красное окрашивание), содержщие ионы магния – хлорофилл (зеленое окрашивание). Витамин В12 (кобалимин) содержит координированный ион кобальта Со2+ в порфириноподобном макроцикле – коррине, состоящем из четырех частично гидрированных пиррольных колец (розовое окрашивание).!}
Ppt%5C34928-slozhnye_belki_ch1_37.jpg" alt=">Chất diệp lục b. Chất diệp lục tham gia vào quá trình quang hợp.">!}
Ppt%5C34928-slozhnye_belki_ch1_38.jpg" alt=">Cytochromes khác nhau về thành phần axit amin của chuỗi peptide, số lượng chuỗi và được chia thành loại a, b,"> Цитохромы различаются аминокислотным составом пептидных цепей, числом цепей и разделяются на типы а, b, с, d. Цитохромы находятся в составе дыхательной цепи и цепи микросомального окисления. Степень окисления железа в составе цитохромов меняется в отличие от гемоглобина и миоглобина Fe2+ ↔ Fe3+!}
Ppt%5C34928-slozhnye_belki_ch1_39.jpg" alt=">Myoglobin (MB) là một loại protein được tìm thấy trong cơ đỏ, chức năng chính của nó là tạo ra chất dự trữ"> Миоглобин (Мв) - белок, находящийся в красных мышцах, основная функция которого - создание запасов О2, необходимых при интенсивной мышечной работе. Мв - сложный белок, содержащий белковую часть - апоМв и небелковую часть - гем. Первичная структура апоМв определяет его компактную глобулярную конформацию и структуру активного центра, к которому присоединяется небелковая часть миоглобина - гем. Кислород, поступающий из крови в мышцы, связывается с Fe2+ гема в составе миоглобина. Мв - мономерный белок, имеющий очень высокое сродство к О2, поэтому отдача кислорода миоглобином происходит только при интенсивной мышечной работе, когда парциальное давление O2 резко снижается. Формирование пространственных структур и функционирование миоглобина.!}
Ppt%5C34928-slozhnye_belki_ch1_40.jpg" alt=">Sự hình thành cấu trúc MV. Ở cơ đỏ, quá trình tổng hợp protein chính xảy ra trên ribosome trong quá trình dịch mã"> Формирование конформации Мв. В красных мышцах на рибосомах в ходе трансляции идет синтез первичной структуры Мв, представленной специфической последовательностью 153 аминокислотных остатков. Вторичная структура Мв содержит восемь α-спиралей, называемых !} bằng chữ Latinh từ A đến H, giữa đó có những vùng không có hình xoắn ốc. Cấu trúc bậc ba của Mb có dạng một cầu nhỏ gọn, trong phần lõm của nó có trung tâm hoạt động nằm giữa các chuỗi xoắn ốc F và E.
Ppt%5C34928-slozhnye_belki_ch1_41.jpg" alt=">Cấu trúc của myoglobin">!}
Ppt%5C34928-slozhnye_belki_ch1_42.jpg" alt=">Đặc điểm cấu trúc và hoạt động của trung tâm hoạt động của Mv. Trung tâm hoạt động của Mv được hình thành chủ yếu bởi các gốc kỵ nước"> Особенности строения и функционирования активного центра Мв. Активный центр Мв сформирован преимущественно гидрофобными радикалами аминокислот, далеко отстоящими друг от друга в первичной структуре (например, Три39 и Фен138). К активному центру присоединяется плохо растворимые в воде лиганды - гем и О2. Гем - специфический лиганд апоМв.!}
Ppt%5C34928-slozhnye_belki_ch1_43.jpg" alt=">Cơ sở của heme được tạo thành từ bốn vòng pyrrole được nối với nhau bằng cầu metyl; ở trung tâm có nguyên tử Fe2+,"> Основу гема составляют четыре пиррольных кольца, соединенных метенильными мостиками; в центре расположен атом Fe2+, соединенный с атомами азота пиррольных колец четырьмя координационными связями. В активном центре Мв кроме гидрофобных радикалов аминокислот имеются также остатки двух аминокислот с гидрофильными радикалами - Гис Е7 (Гис64) и Гис F8 (Гис93).!}
Ppt%5C34928-slozhnye_belki_ch1_44.jpg" alt=">F8 của anh ấy tạo thành liên kết phối hợp với Fe2+ và cố định chắc chắn heme trong trang hoạt động."> Гис F8 образует координационную связь с Fe2+ и прочно фиксирует гем в активном центре. Гис Е7 необходим для правильной ориентации в активном центре другого лиганда - O2 при его взаимодействии с Fe+2 гема. Микроокружение гема создает условия для прочного, но обратимого связывания O2 с Fe+2 и препятствует попаданию в гидрофобный активный центр воды, что может привести к его окислению в Fе3+.!}
Ppt%5C34928-slozhnye_belki_ch1_45.jpg" alt=">Cấu trúc oligomeric của HB và sự điều hòa ái lực HB với O2 bằng phối tử. Huyết sắc tố ở người -"> Олигомерное строение Нв и регуляция сродства Нв к О2 лигандами. Гемоглобины человека - семейство белков, так же как и миоглобин относящиеся к сложным белкам (гемопротеинам). Они имеют тетрамерное строение и содержат две α-цепи, но различаются по строению двух других полипептидных цепей (2α-, 2х-цепи). Строение второй полипептидной цепи определяет особенности функционирования этих форм Нв. Около 98% гемоглобина эритроцитов взрослого человека составляет гемоглобин А (2α-, 2β-цепи). В период внутриутробного развития функционируют два основных типа гемоглобинов: эмбриональный Нв (2α, 2ε), который обнаруживается на ранних этапах развития плода, и гемоглобин F (фетальный) - (2α, 2γ), который приходит на смену раннему гемоглобину плода на шестом месяце внутриутробного развития и только после рождения замещается на Нв А.!}
Ppt%5C34928-slozhnye_belki_ch1_46.jpg" alt=">Hb A là một protein liên quan đến myoglobin (Mb), được tìm thấy trong tế bào hồng cầu của người trưởng thành. Cấu trúc của nó"> Нв А - белок, родственный миоглобину (Мв), содержится в эритроцитах взрослого человека. Строение его отдельных протомеров аналогично таковому у миоглобина. Вторичная и третичная структуры миоглобина и протомеров гемоглобина очень сходны, несмотря на то что в первичной структуре их полипептидных цепей идентичны только 24 аминокислотных остатка (вторичная структура протомеров гемоглобина, так же как миоглобин, содержит восемь α-спиралей, обозначаемых латинскими буквами от А до Н, а третичная структура имеет вид компактной глобулы). Но в отличие от миоглобина гемоглобин имеет олигомерное строение, состоит из четырех полипептидных цепей, соединенных нековалентными связями.!}
Ppt%5C34928-slozhnye_belki_ch1_47.jpg" alt=">Cấu trúc oligomeric của hemoglobin">!}
Ppt%5C34928-slozhnye_belki_ch1_48.jpg" alt=">Mỗi proton Hb được liên kết với một phần không phải protein - heme và các proton lân cận. Sự kết nối của protein"> Каждый протомер Нв связан с небелковой частью - гемом и соседними протомерами. Соединение белковой части Нв с гемом аналогично таковому у миоглобина: в активном центре белка гидрофобные части гема окружены гидрофобными радикалами аминокислот за исключением Гис F8 и Гис Е7, которые расположены по обе стороны от плоскости гема и играют аналогичную роль в функционировании белка и связывании его с кислородом. Кроме того, Гис Е7 выполняет важную дополнительную роль в функционировании Нв. Свободный гем имеет в 25 000 раз более высокое сродство к СО, чем к О2. СО в небольших количествах образуется в организме и, учитывая его высокое сродство к гему, он мог бы нарушать транспорт необходимого для жизни клеток О2. Однако в составе гемоглобина сродство гема к оксиду углерода превышает сродство к О2 всего в 200 раз благодаря наличию в активном центре Гис Е7. Остаток этой аминокислоты создает !} điều kiện tối ưuđể liên kết heme với O2 và làm suy yếu sự tương tác của heme với CO.
Ppt%5C34928-slozhnye_belki_ch1_49.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_50.jpg" alt=">Các vòng pyrrole của heme nằm trong cùng một mặt phẳng và ion Fe2+ ở trạng thái không có oxy Hb"> Пиррольные кольца гема расположены в одной плоскости, а ион Fe2+ в неоксигенированом состоянии Hb выступает над плоскостью на 0,6 А. При присоединении кислорода ион железа погружается в плоскость колец гема. В результате сдвигается и участок полипептидной цепи, нарушаются слабые связи в молекуле Hb и изменяется конформация всей глобулы. Таким образом, присоединение кислорода вызывает изменение пространственной структуры молекулы миоглобина или протомеров гемоглобина.!}
Ppt%5C34928-slozhnye_belki_ch1_51.jpg" alt=">Hemoglobin có thể tồn tại ở cả dạng tự do (deoxyhemoglobin) và dạng oxy hóa, tổng cộng có tới"> Гемоглобин может существовать как в свободной (дезоксигемоглобин), так и в оксигенированной форме, присоединяя до 4 молекул кислорода. Взаимодействие с кислородом 1-го протомера вызывает изменение его конформации, а также кооперативные конформационные изменения остальных протомеров. Сродство к кислороду возрастает, и присоединение О2 к активному центру 2-го протомера происходит легче, вызывая дальнейшую конформационную перестройку всей молекулы. В результате еще сильнее изменяется структура оставшихся протомеров и их активных центров, взаимодействие с О2 еще больше облегчается. В итоге 4-я молекула кислорода присоединяется к Hb примерно в 300 раз легче, чем 1-я. Так происходит в легких при высоком парциальном давлении кислорода.!}
Ppt%5C34928-slozhnye_belki_ch1_52.jpg" alt=">Sự thay đổi hợp tác về hình dạng của phân tử hemoglobin khi tương tác với oxy">!}
Ppt%5C34928-slozhnye_belki_ch1_53.jpg" alt=">Trong các mô có hàm lượng oxy thấp hơn, ngược lại, sự phân tách của từng phân tử O2 tạo điều kiện cho việc giải phóng các phân tử tiếp theo."> В тканях, где содержание кислорода ниже, наоборот, отщепление каждой молекулы О2 облегчает освобождение последующих. Таким образом, взаимодействие олигомерного белка гемоглобина с лигандом (О2) в одном центре связывания приводит к изменению конформации всей молекулы и других, пространственно удаленных центров, расположенных на других субъединицах (принцип «домино»). Подобные взаимосвязанные изменения структуры белка называют кооперативными конформационными изменениями. Они характерны для всех олигомерных белков и используются для регуляции их активности.!}
Ppt%5C34928-slozhnye_belki_ch1_54.jpg" alt=">Sự tương tác của cả hai protein (Mb và Hb) với oxy phụ thuộc vào áp suất riêng phần của nó trong"> Взаимодействие обоих белков (Mb и Hb) с кислородом зависит от его парциального давления в тканях. Эта зависимость имеет разный характер, что связано с их особенностями структуры и функционирования. Гемоглобин имеет S-образную кривую насыщения, которая показывает, что субъединицы белка работают кооперативно, и чем больше кислорода они отдают, тем легче идет освобождение остальных молекул О2. Этот процесс зависит от изменения парциального давления кислорода в тканях. График насыщения миоглобина кислородом имеет характер простой гиперболы, т.е. насыщение Mb кислородом происходит быстро и отражает его функцию - обратимое связывание с кислородом, высвобождаемым гемоглобином, и освобождение в случае интенсивной физической нагрузки.!}
Ppt%5C34928-slozhnye_belki_ch1_55.jpg" alt=">Đường cong bão hòa oxy của Myoglobin và hemoglobin">!}
Ppt%5C34928-slozhnye_belki_ch1_56.jpg" alt=">CO2 và H+, được hình thành trong quá trình dị hóa các chất hữu cơ, làm giảm ái lực của hemoglobin đối với O2 theo tỷ lệ"> CO2 и Н+, образующиеся при катаболизме органических веществ, уменьшают сродство гемоглобина к О2 пропорционально их концентрации. Энергия, необходимая для работы клеток, вырабатывается преимущественно в митохондриях при окислении органических веществ с использованием O2, доставляемого из легких гемоглобином. В результате окисления органических веществ образуются конечные продукты их распада: СО2 и Н2O, количество которых пропорционально интенсивности протекающих процессов окисления. СO2 диффузией попадает из клеток в кровь и проникает в эритроциты, где под действием фермента карбоангидразы превращается в угольную кислоту. Эта слабая кислота диссоциирует на протон и бикарбонат ион. СО2 + Н2О → Н2СО3 → Н+ + НСО3-!}
Ppt%5C34928-slozhnye_belki_ch1_57.jpg" alt=">H+ ion có thể tham gia các gốc His146 trong chuỗi β của hemoglobin, tức là ở các khu vực xa xôi"> Ионы Н+ способны присоединятся к радикалам Гис146 в β-цепях гемоглобина, т.е. в участках, удаленных от гема. Протонирование гемоглобина снижает его сродство к О2, способствует отщеплению О2 от оксиНв, образованию дезоксиНв и увеличивает поступление кислорода в ткани пропорционально количеству образовавшихся протонов. Увеличение количества освобожденного кислорода в зависимости от увеличения концентрации Н+ в эритроцитах называется эффектом Бора (по имени датского физиолога Христиана Бора, впервые открывшего этот эффект). В легких высокое парциальное давление кислорода способствует его связыванию с дезоксиНв, что уменьшает сродство белка к Н+. Освободившиеся протоны под действием карбоангидразы взаимодействуют с бикарбонатами с образованием СО2 и Н2О!}
Ppt%5C34928-slozhnye_belki_ch1_58.jpg" alt=">Sự phụ thuộc ái lực của Hb với O2 vào nồng độ CO2 và proton (hiệu ứng Bohr): A -"> Зависимость сродства Нв к О2 от концентрации СО2 и протонов (эффект Бора): А - влияние концентрации СО2 и Н+ на высвобождение О2 из комплекса с Нв (эффект Бора); Б - оксигенирование дезоксигемоглобина в легких, образование и выделение СО2.!}
Ppt%5C34928-slozhnye_belki_ch1_59.jpg" alt=">CO2 sinh ra đi vào không gian phế nang và được loại bỏ bằng không khí thở ra. Do đó, lượng"> Образовавшийся СО2 поступает в альвеолярное пространство и удаляется с выдыхаемым воздухом. Таким образом, количество высвобождаемого гемоглобином кислорода в тканях регулируется продуктами катаболизма органических веществ: чем интенсивнее распад веществ, например при физических нагрузках, тем выше концентрация СО2 и Н+ и тем больше кислорода получают ткани в результате уменьшения сродства Нв к О2.!}
Ppt%5C34928-slozhnye_belki_ch1_60.jpg" alt=">Sự thay đổi trong hoạt động chức năng của protein khi tương tác với các phối tử khác do thay đổi về hình dạng được gọi là allosteric"> Изменение функциональной активности белка при взаимодействии с другими лигандами вследствие конформационных изменений называется аллостерической регуляцией, а соединения-регуляторы - аллостерическими лигандами или эффекторами. Способность к аллостерической регуляции характерна, как правило, для олигомерных белков, т.е. для проявления аллостерического эффекта необходимо взаимодействие протомеров. При воздействии аллостерических лигандов белки меняют свою конформацию (в том числе и активного центра) и функцию.!}
Ppt%5C34928-slozhnye_belki_ch1_61.jpg" alt=">Sự điều hòa dị lập thể về ái lực của Hb đối với O2 bằng phối tử - 2,3-bis-phosphoglycerate. Trong hồng cầu từ sản phẩm"> Аллостерическая регуляция сродства Нв к О2 лигандом - 2,3-бис-фосфоглицератом. В эритроцитах из продукта окисления глюкозы - 1,3-бисфосфоглицерата синтезируется аллостерический лиганд гемоглобина - 2,3-бисфосфоглицерат (2,3-БФГ). В нормальных условиях концентрация 2,3-БФГ высокая и сравнима с концентрацией Нв. 2,3-БФГ имеет сильный отрицательный заряд (-5).!}
Ppt%5C34928-slozhnye_belki_ch1_62.jpg" alt=">Có một khoang ở trung tâm của phân tử tetrameric hemoglobin. Nó được hình thành bởi dư lượng axit amin của cả bốn proton."> В центре тетрамерной молекулы гемоглобина находится полость. Ее образуют аминокислотные остатки всех четырех протомеров. В капиллярах тканей протонирование Нв (эффект Бора) приводит к разрыву связи между железом гема и О2. В молекуле дезоксигемоглобина по сравнению с оксигемоглобином возникают дополнительные ионные связи, соединяющие протомеры, вследствие чего размеры центральной полости по сравнению с оксигемоглобином увеличиваются. Центральная полость является местом присоединения 2,3-БФГ к гемоглобину. БФГ поступает в полость дезоксигемоглобина. 2,3-БФГ взаимодействует с гемоглобином в участке, удаленном от активных центров белка и относится к аллостерическим (регуляторным) лигандам, а центральная полость Нв является аллостерическим центром. 2,3-БФГ имеет сильный отрицательный заряд и взаимодействует с положительно заряженными группами двух β-цепей Нв. При этом его сродство к О2 снижается в 26 раз. В результате происходит высвобождение кислорода в капиллярах ткани при низком парциальном давлении О2. В легких высокое парциальное давление О2, наоборот, приводит к оксигенированию Нв и освобождению БФГ.!}
Ppt%5C34928-slozhnye_belki_ch1_63.jpg" alt=">Vị trí liên kết BPG nằm trong khoang tích điện dương giữa 4 proton của hemoglobin. Tương tác BPG"> Центр связывания БФГ находится в положительно заряженной полости между 4 протомерами гемоглобина. Взаимодействие БФГ с центром связывания изменяет конформацию α- и β-протомеров НЬ и их активных центров. Сродство НЬ к молекулам О2 снижается и кислород высвобождается в ткани. В легких при высоком парциальном давлении О2 активные центры гемоглобина насыщаются за счет изменения конформации и БФГ вытесняется из аллостерического центра!}
Ppt%5C34928-slozhnye_belki_ch1_64.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_65.jpg" alt=">Do đó, protein oligomeric có các đặc tính mới so với protein đơn phân."> Таким образом, олигомерные белки обладают новыми по сравнению с мономерными белками свойствами. Присоединение лигандов на участках, пространственно удаленных друг от друга (аллостерических), способно вызывать конформационные изменения во всей белковой молекуле. Благодаря взаимодействию с регуляторными лигандами происходит изменение конформации и адаптация функции белковой молекулы к изменениям окружающей среды.!}
Ppt%5C34928-slozhnye_belki_ch1_66.jpg" alt=">Khoảng 15% lượng carbon dioxide có trong máu được vận chuyển bởi các phân tử hemoglobin. Trong các mô, một số phân tử"> Около 15% углекислого газа, присутствующего в крови, переносится молекулами гемоглобина. В тканях часть молекул углекислого газа может присоединится к каждому протомеру молекулы гемоглобина, при этом снижается сродство Hb к кислороду. В легких, наоборот, из-за высокого парциального давления кислорода, О2 связывается с Hb, а СО2 высвобождается.!}
Ppt%5C34928-slozhnye_belki_ch1_67.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_68.jpg" alt=">Trong phân tử hemoglobin S (được gọi là hemoglobin bất thường), 2 chuỗi β hóa ra là đột biến, trong đó"> В молекуле гемоглобина S (так назван аномальный гемоглобин) мутантными оказались 2 β-цепи, в которых глутамат, высокополярная отрицательно заряженная аминокислота в положении 6 была заменена валином, содержащим гидрофобный радикал.!}
Cấu trúc mô-đun | Chủ đề |
Đơn vị mô-đun 1 | 1.1. Tổ chức cấu trúc của protein. Các giai đoạn hình thành cấu trúc tự nhiên của protein 1.2. Cơ bản về hoạt động của protein. Thuốc là phối tử ảnh hưởng đến chức năng protein 1.3. Sự biến tính của protein và khả năng phục hồi tự phát của chúng |
Đơn vị mô-đun 2 | 1.4. Đặc điểm cấu trúc và chức năng của protein oligomeric sử dụng ví dụ về huyết sắc tố 1.5. Duy trì cấu trúc protein tự nhiên trong điều kiện tế bào 1.6. Sự đa dạng của protein. Họ protein sử dụng ví dụ về globulin miễn dịch 1.7. Tính chất hóa lý của protein và phương pháp tách chúng |
Đơn vị mô đun 1 TỔ CHỨC CẤU TRÚC CỦA ĐẠM MONOME VÀ CƠ SỞ CHỨC NĂNG CỦA CHÚNG
Mục tiêu học tập Có khả năng:
1. Sử dụng kiến thức về đặc điểm cấu trúc của protein và sự phụ thuộc của chức năng protein vào cấu trúc của chúng để hiểu cơ chế phát triển các bệnh lý protein di truyền và mắc phải.
2. Giải thích cơ chế hiệu quả điều trị một số loại thuốc là phối tử tương tác với protein và thay đổi hoạt động của chúng.
3. Sử dụng kiến thức về cấu trúc và tính không ổn định về hình dạng của protein để hiểu tính không ổn định về cấu trúc và chức năng cũng như xu hướng biến tính của chúng trong các điều kiện thay đổi.
4. Giải thích việc sử dụng các chất biến tính làm phương tiện khử trùng vật liệu và dụng cụ y tế cũng như chất khử trùng.
Biết:
1. Các cấp độ tổ chức cấu trúc của protein.
2. Tầm quan trọng của cấu trúc bậc một của protein quyết định sự đa dạng về cấu trúc và chức năng của chúng.
3. Cơ chế hình thành trung tâm hoạt động trong protein và sự tương tác cụ thể của nó với phối tử làm nền tảng cho hoạt động của protein.
4. Ví dụ về ảnh hưởng của các phối tử ngoại sinh (thuốc, chất độc, chất độc) đến cấu tạo và hoạt động chức năng của protein.
5. Nguyên nhân, hậu quả biến tính protein, các yếu tố gây biến tính.
6. Ví dụ về việc sử dụng các yếu tố biến tính trong y học làm chất khử trùng và phương tiện khử trùng dụng cụ y tế.
CHỦ ĐỀ 1.1. TỔ CHỨC CẤU TRÚC CỦA PROTEIN. CÁC GIAI ĐOẠN HÌNH THÀNH NGƯỜI BẢN XỨ
CẤU HÌNH PROTEIN
Protein là các phân tử polymer có monome chỉ gồm 20 axit amin-α. Tập hợp và thứ tự kết hợp các axit amin trong protein được xác định bởi cấu trúc gen trong DNA của từng cá thể. Mỗi protein, theo cấu trúc cụ thể của nó, thực hiện chức năng riêng của nó. Tập hợp các protein của một sinh vật nhất định xác định các đặc điểm kiểu hình của nó, cũng như sự hiện diện của các bệnh di truyền hoặc khuynh hướng phát triển của chúng.
1. Axit amin tạo nên protein. Liên kết peptit. Protein là các polyme được tạo thành từ các monome - 20 axit α-amino, công thức chung là
Các axit amin khác nhau về cấu trúc, kích thước và tính chất hóa lý của các gốc gắn với nguyên tử α-carbon. Các nhóm chức năng của axit amin xác định đặc điểm tính chất của các axit amin-α khác nhau. Các gốc được tìm thấy trong axit α-amino có thể được chia thành nhiều nhóm:
Proline, Không giống như 19 monome protein khác, nó không phải là axit amin mà là axit imino; gốc trong proline liên kết với cả nguyên tử α-carbon và nhóm imino
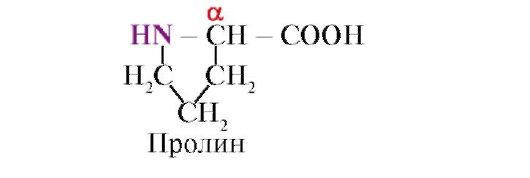 Axit amin khác nhau về độ hòa tan trong nước.Điều này là do khả năng của các gốc tự do tương tác với nước (hydrat).
Axit amin khác nhau về độ hòa tan trong nước.Điều này là do khả năng của các gốc tự do tương tác với nước (hydrat).
ĐẾN ưa nước bao gồm các gốc chứa các nhóm chức anion, cation và không tích điện phân cực.
ĐẾN kỵ nước bao gồm các gốc chứa nhóm metyl, chuỗi hoặc vòng béo.
2. Liên kết peptit nối các axit amin tạo thành peptit. Trong quá trình tổng hợp peptit, nhóm α-carboxyl của axit amin này tương tác với nhóm α-amino của axit amin khác để tạo thành liên kết peptit:
Protein là các polypeptide, tức là các polyme tuyến tính của các axit α-amino được nối với nhau bằng liên kết peptide (Hình 1.1.)
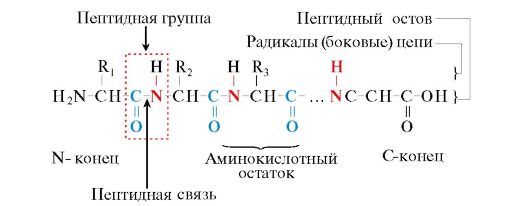 Cơm. 1.1. Thuật ngữ dùng để mô tả cấu trúc của peptit
Cơm. 1.1. Thuật ngữ dùng để mô tả cấu trúc của peptit
Các monome của axit amin tạo nên polypeptide được gọi là dư lượng axit amin. Một chuỗi các nhóm lặp lại - NH-CH-CO- các hình thức xương sống peptit. Dư lượng axit amin có nhóm α-amino tự do được gọi là đầu N và dư lượng có nhóm α-carboxyl tự do được gọi là đầu C. Các peptide được viết và đọc từ đầu N đến đầu C.
Liên kết peptit được hình thành bởi nhóm imino của proline khác với các liên kết peptit khác: nguyên tử nitơ của nhóm peptit thiếu hydro,
thay vào đó, có một liên kết với một gốc, do đó một bên của vòng được bao gồm trong xương sống peptide:
 Các peptide khác nhau về thành phần axit amin, số lượng axit amin và thứ tự liên kết axit amin, ví dụ Ser-Ala-Glu-Gis và His-Glu-Ala-Ser là hai peptide khác nhau.
Các peptide khác nhau về thành phần axit amin, số lượng axit amin và thứ tự liên kết axit amin, ví dụ Ser-Ala-Glu-Gis và His-Glu-Ala-Ser là hai peptide khác nhau.
Liên kết peptide rất mạnh và quá trình thủy phân hóa học không dùng enzyme của chúng đòi hỏi các điều kiện khắc nghiệt: protein được phân tích được thủy phân trong axit clohydric đậm đặc ở nhiệt độ khoảng 110° trong 24 giờ. Trong tế bào sống, liên kết peptide có thể bị phá vỡ bởi enzyme phân giải protein, gọi điện protease hoặc hydrolase peptide.
3. Cấu trúc bậc một của protein. Dư lượng axit amin trong chuỗi peptide của các protein khác nhau không xen kẽ một cách ngẫu nhiên mà được sắp xếp theo một trật tự nhất định. Trình tự tuyến tính hoặc thứ tự xen kẽ của các gốc axit amin trong chuỗi polypeptide được gọi là cấu trúc bậc một của protein.
Cấu trúc bậc một của mỗi protein riêng lẻ được mã hóa trong phân tử DNA (trong vùng gọi là gen) và được hiện thực hóa trong quá trình phiên mã (sao chép thông tin vào mRNA) và dịch mã (tổng hợp cấu trúc bậc một của protein). Do đó, cấu trúc chính của protein của một cá nhân là thông tin được truyền từ cha mẹ sang con cái, thông tin này xác định các đặc điểm cấu trúc của protein của một sinh vật nhất định, mà chức năng của các protein hiện có phụ thuộc vào đó (Hình 1.2.).
 Cơm. 1.2. Mối quan hệ giữa kiểu gen và cấu tạo của các protein được tổng hợp trong cơ thể con người
Cơm. 1.2. Mối quan hệ giữa kiểu gen và cấu tạo của các protein được tổng hợp trong cơ thể con người
Mỗi loại trong số khoảng 100.000 protein riêng lẻ trong cơ thể con người có độc nhất cấu trúc chính. Các phân tử của cùng một loại protein (ví dụ, albumin) có cùng các gốc axit amin xen kẽ, giúp phân biệt albumin với bất kỳ protein riêng lẻ nào khác.
Trình tự các gốc axit amin trong chuỗi peptide có thể được coi là một dạng ghi lại thông tin. Thông tin này xác định sự sắp xếp không gian của chuỗi peptide tuyến tính thành cấu trúc ba chiều nhỏ gọn hơn gọi là hình dáng sóc. Quá trình hình thành cấu trúc protein có chức năng hoạt động được gọi là gấp
4. Cấu hình protein. Có thể xoay tự do trong khung peptide giữa nguyên tử nitơ của nhóm peptide và nguyên tử α-carbon lân cận, cũng như giữa nguyên tử α-carbon và carbon của nhóm carbonyl. Do sự tương tác của các nhóm chức năng của dư lượng axit amin, cấu trúc bậc một của protein có thể có cấu trúc không gian phức tạp hơn. Trong protein hình cầu, có hai mức độ gấp nếp chính của chuỗi peptide: sơ trung Và cấu trúc đại học.
Cấu trúc bậc hai của protein là cấu trúc không gian được hình thành do sự hình thành liên kết hydro giữa các nhóm chức -C=O và -NH- của xương sống peptide. Trong trường hợp này, chuỗi peptide có thể có cấu trúc đều đặn gồm hai loại: xoắn ốc α Và cấu trúc β.
TRONG xoắn ốc α liên kết hydro được hình thành giữa nguyên tử oxy của nhóm carbonyl và hydro của nitơ amit của axit amin thứ 4 từ nó; chuỗi bên của dư lượng axit amin
nằm dọc theo ngoại vi của đường xoắn ốc, không tham gia vào quá trình hình thành cấu trúc thứ cấp (Hình 1.3.).
Các gốc tự do số lượng lớn, hoặc các gốc mang điện tích bằng nhau, ngăn cản sự hình thành chuỗi xoắn ốc. Dư lượng proline, có cấu trúc vòng, làm gián đoạn chuỗi xoắn α, do thiếu hydro ở nguyên tử nitơ trong chuỗi peptide nên không thể hình thành liên kết hydro. Liên kết giữa nitơ và nguyên tử α-carbon là một phần của vòng proline, do đó xương sống peptide bị uốn cong tại điểm này.
Cấu trúc βđược hình thành giữa các vùng tuyến tính của khung peptide của một chuỗi polypeptide, từ đó hình thành các cấu trúc gấp nếp. Chuỗi polypeptide hoặc các bộ phận của nó có thể hình thành song song hoặc cấu trúc β phản song song. Trong trường hợp đầu tiên, đầu N và đầu C của chuỗi peptide tương tác trùng nhau, và trong trường hợp thứ hai, chúng có hướng ngược lại (Hình 1.4).
Cơm. 1.3. Cấu trúc bậc hai của protein - chuỗi xoắn α
 Cơm. 1.4. Cấu trúc tấm β song song và phản song song
Cơm. 1.4. Cấu trúc tấm β song song và phản song song
Cấu trúc β được biểu thị bằng mũi tên rộng: A - Cấu trúc β phản song song. B - Cấu trúc tấm β song song
Ở một số protein, cấu trúc β có thể được hình thành do sự hình thành liên kết hydro giữa các nguyên tử của khung peptide của các chuỗi polypeptide khác nhau.
Cũng được tìm thấy trong protein khu vực có thứ cấp không đều cấu trúc, bao gồm các chỗ uốn cong, vòng lặp và vòng quay của khung polypeptide. Chúng thường nằm ở những nơi mà hướng của chuỗi peptide thay đổi, chẳng hạn như khi hình thành cấu trúc tấm β song song.
Dựa trên sự hiện diện của cấu trúc xoắn α và cấu trúc β, protein hình cầu có thể được chia thành bốn loại.
Cơm. 1.5. Cấu trúc thứ cấp của myoglobin (A) và chuỗi β hemoglobin (B), chứa 8 chuỗi xoắn α

 Cơm. 1.6. Cấu trúc thứ cấp của miền triosephosphate isomerase và pyruvate kinase
Cơm. 1.6. Cấu trúc thứ cấp của miền triosephosphate isomerase và pyruvate kinase
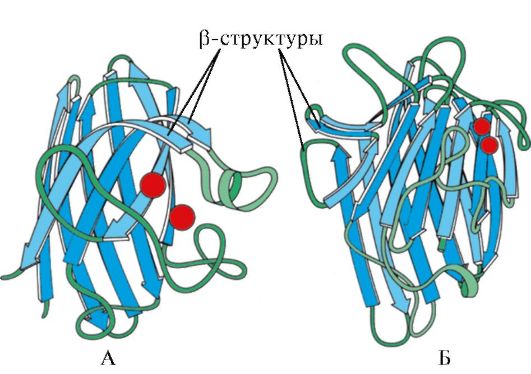 Cơm. 1.7. Cấu trúc thứ cấp của miền cố định của immunoglobulin (A) và enzyme superoxide dismutase (B)
Cơm. 1.7. Cấu trúc thứ cấp của miền cố định của immunoglobulin (A) và enzyme superoxide dismutase (B)
TRONG loại thứ tư bao gồm các protein có chứa một lượng nhỏ cấu trúc thứ cấp thông thường. Những protein này bao gồm các protein nhỏ giàu cysteine hoặc metallicoprotein.
Cấu trúc bậc ba của protein- một loại cấu hình được hình thành do sự tương tác giữa các gốc axit amin, có thể nằm ở một khoảng cách đáng kể với nhau trong chuỗi peptide. Hầu hết các protein tạo thành cấu trúc không gian giống như một khối cầu (protein hình cầu).
Vì các gốc axit amin kỵ nước có xu hướng kết hợp thông qua cái gọi là tương tác kỵ nước và lực van der Waals liên phân tử, một lõi kỵ nước dày đặc được hình thành bên trong giọt protein. Các gốc ion hóa và không ion hóa ưa nước chủ yếu nằm trên bề mặt protein và xác định khả năng hòa tan của nó trong nước.
 Cơm. 1.8. Các loại liên kết phát sinh giữa các gốc axit amin trong quá trình hình thành cấu trúc bậc ba của protein
Cơm. 1.8. Các loại liên kết phát sinh giữa các gốc axit amin trong quá trình hình thành cấu trúc bậc ba của protein
1 - sự gắn kết- xảy ra giữa các nhóm chức năng tích điện dương và âm;
2 - liên kết hydro- xảy ra giữa nhóm không tích điện ưa nước và nhóm ưa nước bất kỳ khác;
3 - tương tác kỵ nước- phát sinh giữa các gốc kỵ nước;
4 - liên kết disulfua- được hình thành do quá trình oxy hóa các nhóm SH của dư lượng cystein và sự tương tác của chúng với nhau
Các gốc axit amin ưa nước nằm bên trong lõi kỵ nước có thể tương tác với nhau bằng cách sử dụng ion Và liên kết hydro(Hình 1.8).
Liên kết ion và hydro, cũng như tương tác kỵ nước, rất yếu: năng lượng của chúng không cao hơn nhiều so với năng lượng chuyển động nhiệt của các phân tử tại nhiệt độ phòng. Cấu hình của protein được duy trì nhờ sự hình thành nhiều liên kết yếu như vậy. Vì các nguyên tử tạo nên protein chuyển động liên tục nên có thể phá vỡ một số liên kết yếu và hình thành các liên kết khác, dẫn đến sự chuyển động nhẹ của từng phần riêng lẻ của chuỗi polypeptide. Đặc tính của protein là thay đổi hình dạng do phá vỡ một số liên kết và hình thành các liên kết yếu khác được gọi là tính không ổn định về hình dạng.
Cơ thể con người có những hệ thống hỗ trợ cân bằng nội môi- sự ổn định của môi trường bên trong trong giới hạn nhất định có thể chấp nhận được đối với một cơ thể khỏe mạnh. Trong điều kiện cân bằng nội môi, những thay đổi nhỏ về hình dạng không phá vỡ cấu trúc và chức năng tổng thể của protein. Cấu hình có chức năng hoạt động của protein được gọi là hình dạng bản địa. Những thay đổi trong môi trường bên trong (ví dụ: nồng độ glucose, ion Ca, proton, v.v.) dẫn đến những thay đổi về cấu trúc và sự phá vỡ các chức năng của protein.
Cấu trúc bậc ba của một số protein được ổn định liên kết disulfua,được hình thành do sự tương tác của nhóm -SH của hai dư lượng
 Cơm. 1.9. Sự hình thành liên kết disulfide trong phân tử protein
Cơm. 1.9. Sự hình thành liên kết disulfide trong phân tử protein
cystein (Hình 1.9). Hầu hết các protein nội bào không có liên kết cộng hóa trị disulfide trong cấu trúc bậc ba của chúng. Sự hiện diện của chúng là đặc trưng của các protein do tế bào tiết ra, đảm bảo tính ổn định cao hơn của chúng trong điều kiện ngoại bào. Do đó, liên kết disulfide hiện diện trong các phân tử insulin và globulin miễn dịch.
insulin- một loại hormone protein được tổng hợp trong tế bào β của tuyến tụy và tiết vào máu để đáp ứng với sự gia tăng nồng độ glucose trong máu. Trong cấu trúc của insulin, có hai liên kết disulfide nối chuỗi polypeptide A và B và một liên kết disulfide trong chuỗi A (Hình 1.10).
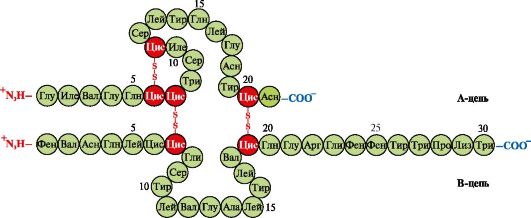 Cơm. 1.10. Liên kết disulfide trong cấu trúc của insulin
Cơm. 1.10. Liên kết disulfide trong cấu trúc của insulin
5. Cấu trúc siêu trung học của protein. Trong các protein có cấu trúc và chức năng bậc một khác nhau, đôi khi chúng được phát hiện sự kết hợp tương tự và vị trí tương đối của các cấu trúc thứ cấp,được gọi là cấu trúc siêu thứ cấp. Nó chiếm vị trí trung gian giữa cấu trúc bậc hai và bậc ba, vì nó là sự kết hợp cụ thể của các yếu tố của cấu trúc bậc hai trong quá trình hình thành cấu trúc bậc ba của protein. Các cấu trúc siêu trung học có các tên cụ thể, chẳng hạn như “α-helix-turn-a-helix”, “dây kéo leucine”, “ngón tay kẽm”, v.v. Những cấu trúc siêu trung học như vậy là đặc trưng của các protein liên kết DNA.
"Dây kéo Leucine." Loại cấu trúc siêu thứ cấp này được sử dụng để nối hai protein lại với nhau. Trên bề mặt của các protein tương tác có các vùng xoắn ốc chứa ít nhất bốn gốc leucine. Dư lượng Leucine trong chuỗi xoắn α nằm cách nhau sáu axit amin. Vì mỗi vòng xoắn α chứa 3,6 gốc axit amin nên các gốc leucine nằm trên bề mặt của mỗi vòng xoắn thứ hai. Dư lượng leucine của chuỗi xoắn α của một protein có thể tương tác với dư lượng leucine của protein khác (tương tác kỵ nước), kết nối chúng lại với nhau (Hình 1.11.). Nhiều protein liên kết DNA hoạt động trong các phức hợp oligomeric trong đó các tiểu đơn vị riêng lẻ được liên kết với nhau bằng “dây kéo leucine”.
 Cơm. 1.11. "Dây kéo Leucine" giữa các vùng xoắn ốc của hai protein
Cơm. 1.11. "Dây kéo Leucine" giữa các vùng xoắn ốc của hai protein
Một ví dụ về các protein như vậy là histone. Histones- protein hạt nhân, chứa một số lượng lớn axit amin tích điện dương - arginine và lysine (lên tới 80%). Các phân tử histone được kết hợp thành các phức hợp oligomeric chứa tám monome sử dụng “dây kéo leucine”, mặc dù các phân tử này có điện tích đồng âm đáng kể.
"ngón tay kẽm"- một biến thể của cấu trúc siêu thứ cấp, đặc trưng của protein gắn DNA, có dạng mảnh dài trên bề mặt protein và chứa khoảng 20 gốc axit amin (Hình 1.12). Hình dạng “ngón tay mở rộng” được hỗ trợ bởi một nguyên tử kẽm liên kết với bốn gốc axit amin - hai dư lượng cysteine và hai dư lượng histidine. Trong một số trường hợp, thay vì dư lượng histidine lại có dư lượng cystein. Hai dư lượng cystein nằm gần nhau được tách ra khỏi hai dư lượng Gisili còn lại bằng chuỗi Cys gồm khoảng 12 dư lượng axit amin. Vùng protein này tạo thành một chuỗi xoắn α, các gốc của nó có thể liên kết đặc hiệu với các vùng điều hòa của rãnh chính của DNA. Tính đặc hiệu ràng buộc riêng lẻ
 Cơm. 1.12. Cấu trúc chính của vùng protein liên kết DNA tạo thành cấu trúc “ngón tay kẽm” (các chữ cái chỉ các axit amin tạo nên cấu trúc này)
Cơm. 1.12. Cấu trúc chính của vùng protein liên kết DNA tạo thành cấu trúc “ngón tay kẽm” (các chữ cái chỉ các axit amin tạo nên cấu trúc này)
Protein liên kết DNA điều hòa phụ thuộc vào trình tự các gốc axit amin nằm ở vùng ngón tay kẽm. Đặc biệt, những cấu trúc như vậy chứa các thụ thể hormone steroid liên quan đến việc điều hòa phiên mã (đọc thông tin từ DNA sang RNA).
CHỦ ĐỀ 1.2. CƠ SỞ VỀ CHỨC NĂNG CỦA PROTEIN. THUỐC NHƯ LIGAND ẢNH HƯỞNG CHỨC NĂNG PROTEIN
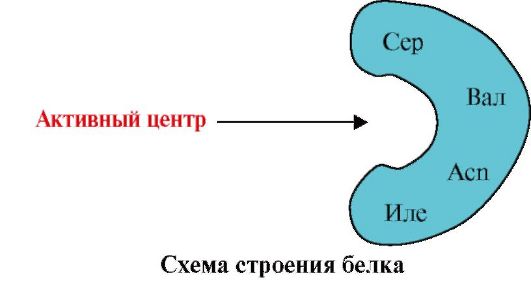
1. Trung tâm hoạt động của protein và sự tương tác của nó với phối tử. Trong quá trình hình thành cấu trúc bậc ba, một vùng được hình thành trên bề mặt của protein hoạt động chức năng, thường ở dạng lõm, được hình thành bởi các gốc axit amin cách xa nhau trong cấu trúc bậc một. Vùng này có cấu trúc duy nhất cho một protein nhất định và có khả năng tương tác đặc hiệu với một phân tử cụ thể hoặc nhóm các phân tử tương tự, được gọi là vị trí liên kết phối tử protein hoặc vị trí hoạt động. Phối tử là các phân tử tương tác với protein.
Độ đặc hiệu cao Sự tương tác của protein với phối tử được đảm bảo bởi sự bổ sung giữa cấu trúc của trung tâm hoạt động với cấu trúc của phối tử.
Tính bổ sung- đây là sự tương ứng về không gian và hóa học của các bề mặt tương tác. Trung tâm hoạt động không chỉ phải tương ứng về mặt không gian với phối tử có trong nó mà còn phải hình thành các liên kết (tương tác ion, hydro và kỵ nước) giữa các nhóm chức của các gốc có trong trung tâm hoạt động và phối tử giữ phối tử. ở trung tâm hoạt động (Hình 1.13).
 Cơm. 1.13. Tương tác bổ sung của protein với phối tử
Cơm. 1.13. Tương tác bổ sung của protein với phối tử
Một số phối tử, khi gắn vào trung tâm hoạt động của protein, đóng vai trò phụ trợ trong hoạt động của protein. Các phối tử như vậy được gọi là đồng yếu tố và các protein có chứa phần không phải protein được gọi là protein phức tạp(trái ngược với các protein đơn giản, chỉ bao gồm phần protein). Phần không phải protein, gắn chặt với protein, được gọi là nhóm giả. Ví dụ, myoglobin, hemoglobin và cytochromes chứa một nhóm giả, heme, chứa ion sắt, gắn chặt vào trung tâm hoạt động. Các protein phức tạp chứa heme được gọi là hemoprotein.
Khi các phối tử cụ thể được gắn vào protein, chức năng của các protein này sẽ được thể hiện. Như vậy, albumin, protein quan trọng nhất trong huyết tương, thể hiện chức năng vận chuyển bằng cách gắn các phối tử kỵ nước như axit béo, bilirubin, một số thuốc, v.v. vào trung tâm hoạt động (Hình 1.14)
Các phối tử tương tác với cấu trúc ba chiều của chuỗi peptide không chỉ có thể là các phân tử hữu cơ và vô cơ phân tử thấp mà còn có thể là các đại phân tử:
DNA (ví dụ với các protein liên kết với DNA đã thảo luận ở trên);
Polysacarit;
 Cơm. 1.14. Mối quan hệ giữa kiểu gen và kiểu hình
Cơm. 1.14. Mối quan hệ giữa kiểu gen và kiểu hình
Cấu trúc bậc một duy nhất của protein ở người, được mã hóa trong phân tử DNA, được hiện thực hóa trong các tế bào dưới dạng hình dạng, cấu trúc trung tâm hoạt động và chức năng protein độc đáo
Trong những trường hợp này, protein nhận biết một vùng cụ thể của phối tử tương xứng và bổ sung cho vị trí gắn kết. Như vậy, trên bề mặt tế bào gan có các protein thụ thể của hormone insulin, loại hormone này cũng có cấu trúc protein. Sự tương tác của insulin với thụ thể gây ra sự thay đổi về hình dạng và kích hoạt hệ thống tín hiệu, dẫn đến việc lưu trữ chất dinh dưỡng trong tế bào gan sau bữa ăn.
Như vậy, Hoạt động của protein dựa trên sự tương tác cụ thể của trung tâm hoạt động của protein với phối tử.
2. Cấu trúc miền và vai trò của nó trong hoạt động của protein. Chuỗi polypeptide dài của protein hình cầu thường gấp lại thành nhiều vùng nhỏ gọn, tương đối độc lập. Chúng có cấu trúc bậc ba độc lập, gợi nhớ đến cấu trúc của protein hình cầu và được gọi là tên miền. Nhờ cấu trúc miền của protein, cấu trúc bậc ba của chúng dễ hình thành hơn.
Trong protein miền, các vị trí liên kết phối tử thường nằm giữa các miền. Vì vậy, trypsin là một enzyme phân giải protein được sản xuất bởi phần ngoại tiết của tuyến tụy và cần thiết cho quá trình tiêu hóa protein thực phẩm. Nó có cấu trúc hai miền và trung tâm liên kết của trypsin với phối tử của nó - protein thực phẩm - nằm ở rãnh giữa hai miền. Ở trung tâm hoạt động, các điều kiện cần thiết để tạo ra sự liên kết hiệu quả của một vị trí cụ thể của protein thực phẩm và quá trình thủy phân các liên kết peptide của nó.
Các miền khác nhau trong protein có thể di chuyển tương đối với nhau khi trung tâm hoạt động tương tác với phối tử (Hình 1.15).
Hexokinase- một loại enzyme xúc tác quá trình phosphoryl hóa glucose bằng ATP. Vị trí hoạt động của enzyme nằm ở khe hở giữa hai miền. Khi hexokinase liên kết với glucose, các miền xung quanh nó đóng lại và chất nền bị giữ lại, nơi xảy ra quá trình phosphoryl hóa (xem Hình 1.15).
 Cơm. 1.15. Liên kết các miền hexokinase với glucose
Cơm. 1.15. Liên kết các miền hexokinase với glucose
Ở một số protein, các miền thực hiện các chức năng độc lập bằng cách liên kết với các phối tử khác nhau. Những protein như vậy được gọi là đa chức năng.
3. Thuốc là phối tử có tác dụng tác động lên chức năng của protein. Sự tương tác của protein với phối tử là cụ thể. Tuy nhiên, do tính không ổn định về hình dạng của protein và trung tâm hoạt động của nó, nên có thể chọn một chất khác cũng có thể tương tác với protein ở trung tâm hoạt động hoặc phần khác của phân tử.
Chất có cấu trúc tương tự phối tử tự nhiên được gọi là cấu trúc tương tự của phối tử hoặc phối tử không tự nhiên. Nó cũng tương tác với protein tại vị trí hoạt động. Một chất có cấu trúc tương tự của phối tử có thể vừa tăng cường chức năng của protein (chất chủ vận), và giảm nó (nhân vật phản diện). Phối tử và các chất tương tự có cấu trúc của nó cạnh tranh với nhau để liên kết với protein ở cùng một vị trí. Những chất như vậy được gọi là bộ điều biến cạnh tranh(bộ điều chỉnh) chức năng của protein. Nhiều loại thuốc hoạt động như chất ức chế protein. Một số trong số chúng thu được bằng cách biến đổi hóa học các phối tử tự nhiên. Chất ức chế chức năng protein có thể là thuốc và chất độc.
Atropine là chất ức chế cạnh tranh của thụ thể M-cholinergic. Acetylcholine là chất dẫn truyền thần kinh để truyền các xung thần kinh thông qua các khớp thần kinh cholinergic. Để thực hiện sự kích thích, acetylcholine được giải phóng vào khe hở tiếp hợp phải tương tác với protein thụ thể của màng sau synap. Hai loại được tìm thấy Thụ thể cholinergic:
Thụ thể M Ngoài acetylcholine, nó còn tương tác có chọn lọc với muscarine (độc tố của ruồi giấm). Các thụ thể M - cholinergic hiện diện trên cơ trơn và khi tương tác với acetylcholine sẽ gây ra sự co bóp của chúng;
thụ thể H liên kết đặc biệt với nicotin. Các thụ thể N-cholinergic được tìm thấy ở các khớp thần kinh của cơ vân.
Chất ức chế đặc hiệu Thụ thể M-cholinergic là atropin. Nó được tìm thấy trong cây belladonna và henbane.
 Atropine có các nhóm chức năng có cấu trúc tương tự acetylcholine và sự sắp xếp không gian của chúng, do đó nó là chất ức chế cạnh tranh của thụ thể M-cholinergic. Xét rằng sự gắn kết của acetylcholine với các thụ thể M-cholinergic gây ra sự co cơ trơn, atropine được sử dụng như một loại thuốc làm giảm sự co thắt của chúng. (chống co thắt). Vì vậy, người ta biết sử dụng atropine để thư giãn cơ mắt khi quan sát đáy mắt, cũng như để giảm co thắt khi bị đau bụng tiêu hóa. Các thụ thể M-cholinergic cũng có mặt ở trung tâm hệ thần kinh(CNS), do đó, liều lượng lớn atropine có thể gây ra phản ứng không mong muốn từ hệ thần kinh trung ương: kích động vận động và tinh thần, ảo giác, co giật.
Atropine có các nhóm chức năng có cấu trúc tương tự acetylcholine và sự sắp xếp không gian của chúng, do đó nó là chất ức chế cạnh tranh của thụ thể M-cholinergic. Xét rằng sự gắn kết của acetylcholine với các thụ thể M-cholinergic gây ra sự co cơ trơn, atropine được sử dụng như một loại thuốc làm giảm sự co thắt của chúng. (chống co thắt). Vì vậy, người ta biết sử dụng atropine để thư giãn cơ mắt khi quan sát đáy mắt, cũng như để giảm co thắt khi bị đau bụng tiêu hóa. Các thụ thể M-cholinergic cũng có mặt ở trung tâm hệ thần kinh(CNS), do đó, liều lượng lớn atropine có thể gây ra phản ứng không mong muốn từ hệ thần kinh trung ương: kích động vận động và tinh thần, ảo giác, co giật.
Ditilin là chất chủ vận cạnh tranh của thụ thể H-cholinergic, ức chế chức năng của các khớp thần kinh cơ.
 Các khớp thần kinh cơ của cơ xương có chứa thụ thể H-cholinergic. Sự tương tác của chúng với acetylcholine dẫn đến sự co cơ. Trong một số hoạt động phẫu thuật, cũng như trong nghiên cứu nội soi, các loại thuốc được sử dụng có tác dụng làm giãn cơ xương. (thuốc giãn cơ). Chúng bao gồm dithiline, một chất có cấu trúc tương tự acetylcholine. Nó gắn vào các thụ thể H-cholinergic, nhưng không giống như acetylcholine, nó bị enzyme acetylcholinesterase phá hủy rất chậm. Do việc mở các kênh ion kéo dài và quá trình khử cực kéo dài của màng, quá trình dẫn truyền các xung thần kinh bị gián đoạn và xảy ra hiện tượng giãn cơ. Ban đầu, những đặc tính này được phát hiện trong chất độc curare, đó là lý do tại sao những loại thuốc này được gọi là giống curare.
Các khớp thần kinh cơ của cơ xương có chứa thụ thể H-cholinergic. Sự tương tác của chúng với acetylcholine dẫn đến sự co cơ. Trong một số hoạt động phẫu thuật, cũng như trong nghiên cứu nội soi, các loại thuốc được sử dụng có tác dụng làm giãn cơ xương. (thuốc giãn cơ). Chúng bao gồm dithiline, một chất có cấu trúc tương tự acetylcholine. Nó gắn vào các thụ thể H-cholinergic, nhưng không giống như acetylcholine, nó bị enzyme acetylcholinesterase phá hủy rất chậm. Do việc mở các kênh ion kéo dài và quá trình khử cực kéo dài của màng, quá trình dẫn truyền các xung thần kinh bị gián đoạn và xảy ra hiện tượng giãn cơ. Ban đầu, những đặc tính này được phát hiện trong chất độc curare, đó là lý do tại sao những loại thuốc này được gọi là giống curare.
CHỦ ĐỀ 1.3. BIẾN ĐỔI PROTEIN VÀ KHẢ NĂNG PHỤC HỒI TỰ NHIÊN CỦA CHÚNG
1. Do cấu hình tự nhiên của protein được duy trì do tương tác yếu nên những thay đổi về thành phần và tính chất của môi trường xung quanh protein, việc tiếp xúc với thuốc thử hóa học và các yếu tố vật lý gây ra sự thay đổi về cấu hình của chúng (đặc tính của khả năng hình dạng). Phá vỡ một số lượng lớn liên kết dẫn đến phá hủy cấu trúc tự nhiên và biến tính protein.
Sự biến tính của protein- đây là sự phá hủy cấu trúc tự nhiên của chúng dưới tác động của các tác nhân biến tính, gây ra bởi sự đứt gãy các liên kết yếu giúp ổn định cấu trúc không gian của protein. Sự biến tính đi kèm với sự phá hủy cấu trúc ba chiều độc đáo và trung tâm hoạt động của protein cũng như làm mất hoạt tính sinh học của nó (Hình 1.16).
Tất cả các phân tử bị biến tính của một protein đều có cấu hình ngẫu nhiên khác với các phân tử khác của cùng một loại protein. Các gốc axit amin hình thành nên trung tâm hoạt động hóa ra lại cách xa nhau về mặt không gian, tức là vị trí liên kết cụ thể của protein với phối tử bị phá hủy. Trong quá trình biến tính, cấu trúc chính của protein không thay đổi.
Ứng dụng chất biến tính trong nghiên cứu sinh học và y học. Trong các nghiên cứu sinh hóa, trước khi xác định các hợp chất có trọng lượng phân tử thấp trong vật liệu sinh học, protein thường được loại bỏ khỏi dung dịch trước tiên. Axit trichloroacetic (TCA) thường được sử dụng cho mục đích này. Sau khi thêm TCA vào dung dịch, các protein bị biến tính sẽ kết tủa và dễ dàng loại bỏ bằng cách lọc (Bảng 1.1.)
Trong y học, chất biến tính thường được dùng để khử trùng các dụng cụ, vật liệu y tế trong nồi hấp (chất biến tính là nhiệt độ cao) và làm chất khử trùng (cồn, phenol, cloramin) để xử lý các bề mặt bị ô nhiễm có chứa hệ vi sinh vật gây bệnh.
2. Tái kích hoạt protein tự phát- bằng chứng về tính quyết định của cấu trúc bậc một, hình dạng và chức năng của protein. Các protein riêng lẻ là sản phẩm của một gen có trình tự axit amin giống hệt nhau và có cùng cấu hình trong tế bào. Kết luận cơ bản rằng cấu trúc sơ cấp của protein đã chứa thông tin về cấu trúc và chức năng của nó được đưa ra trên cơ sở khả năng của một số protein (đặc biệt là ribonuclease và myoglobin) có thể tự phục hồi - khôi phục cấu trúc tự nhiên của chúng sau khi biến tính.
Sự hình thành cấu trúc protein không gian được thực hiện bằng phương pháp tự lắp ráp - một quá trình tự phát trong đó chuỗi polypeptide, có cấu trúc bậc một duy nhất, có xu hướng chấp nhận cấu hình có năng lượng tự do thấp nhất trong dung dịch. Khả năng phục hồi các protein giữ được cấu trúc bậc một sau khi biến tính đã được mô tả trong một thí nghiệm với enzyme ribonuclease.
Ribonuclease là một enzyme phá vỡ liên kết giữa các nucleotide riêng lẻ trong phân tử RNA. Protein hình cầu này có một chuỗi polypeptide, cấu trúc cấp ba được ổn định bởi nhiều liên kết yếu và bốn liên kết disulfide.
Xử lý ribonuclease bằng urê, chất phá vỡ liên kết hydro trong phân tử và chất khử, phá vỡ liên kết disulfide, dẫn đến biến tính enzyme và mất hoạt động của nó.
Việc loại bỏ các tác nhân gây biến tính bằng phương pháp lọc máu sẽ dẫn đến sự phục hồi cấu trúc và chức năng của protein, tức là để tái sinh. (Hình 1.17).
 Cơm. 1.17. Biến tính và phục hồi ribonuclease
Cơm. 1.17. Biến tính và phục hồi ribonuclease
A - cấu trúc tự nhiên của ribonuclease, trong cấu trúc bậc ba có bốn liên kết disulfide; B - phân tử ribonuclease bị biến tính;
B - phân tử ribonuclease được kích hoạt lại với cấu trúc và chức năng được phục hồi
1. Điền vào bảng 1.2.
Bảng 1.2. Phân loại axit amin theo độ phân cực của gốc tự do
2. Viết công thức của tetrapeptit:
Asp - Pro - Fen - Liz
a) nêu rõ các nhóm lặp lại trong peptit tạo nên khung peptit và các nhóm biến đổi được đại diện bởi các gốc axit amin;
b) dán nhãn đầu N và đầu C;
c) đánh dấu các liên kết peptit;
d) viết một peptit khác gồm các axit amin giống nhau;
e) đếm số biến thể có thể có của tetrapeptit có thành phần axit amin tương tự.
3. Giải thích vai trò của cấu trúc bậc một của protein bằng ví dụ phân tích so sánh hai loại hormone peptide có cấu trúc tương tự và gần gũi về mặt tiến hóa của tuyến yên thần kinh ở động vật có vú - oxytocin và vasopressin (Bảng 1.3).
Bảng 1.3. Cấu trúc và chức năng của oxytocin và vasopressin
 Đối với điều này:
Đối với điều này:
a) so sánh thành phần và trình tự axit amin của hai peptit;
b) tìm sự giống nhau về cấu trúc bậc một của hai peptit và sự giống nhau về tác dụng sinh học của chúng;
c) tìm sự khác biệt về cấu trúc của hai peptit và sự khác biệt về chức năng của chúng;
d) rút ra kết luận về ảnh hưởng của cấu trúc bậc một của peptit đến chức năng của chúng.
4. Mô tả các giai đoạn chính của sự hình thành cấu trúc của protein hình cầu (cấu trúc bậc hai, bậc ba, khái niệm cấu trúc siêu bậc). Nêu các loại liên kết tham gia hình thành cấu trúc protein. Những gốc axit amin nào có thể tham gia hình thành các tương tác kỵ nước, liên kết ion, hydro.
Cho ví dụ.
5. Xác định khái niệm “khả năng hình thành của protein”, chỉ ra lý do tồn tại và ý nghĩa của nó.
6. Mở rộng nghĩa của cụm từ sau: “Hoạt động của protein dựa trên sự tương tác cụ thể của chúng với phối tử”, sử dụng các thuật ngữ và giải thích ý nghĩa của chúng: cấu tạo protein, trung tâm hoạt động, phối tử, tính bổ sung, chức năng protein.
7. Sử dụng một ví dụ, giải thích các miền là gì và vai trò của chúng trong hoạt động của protein.
NHIỆM VỤ TỰ KIỂM SOÁT
1. Cuộc thi đấu.
Nhóm chức trong gốc axit amin:
A. Nhóm carboxyl B. Nhóm hydroxyl C Nhóm guanidine D. Nhóm Thiol E. Nhóm amino
2. Chọn những đáp án đúng.
Axit amin có gốc không tích điện phân cực là:
A. Cis B. Asn
B. Glam G. Ba
3. Chọn những đáp án đúng.
Các gốc axit amin:
A. Cung cấp tính đặc hiệu của cấu trúc bậc một B. Tham gia vào việc hình thành cấu trúc bậc ba
B. Nằm trên bề mặt của protein, chúng ảnh hưởng đến độ hòa tan của protein D. Hình thành trung tâm hoạt động
D. Tham gia hình thành liên kết peptit
4. Chọn những đáp án đúng.
Tương tác kỵ nước có thể hình thành giữa các gốc axit amin:
A. Tre Lay B. Pro Ba
B. Met Ile G. Tir Ala D. Val Fen
5. Chọn những đáp án đúng.
Liên kết ion có thể hình thành giữa các gốc axit amin:
A. Gln Asp B. Apr Liz
B. Liz Glam G. Gis Asp D. Asn Tháng Tư
6. Chọn những đáp án đúng.
Liên kết hydro có thể hình thành giữa các gốc axit amin:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Cuộc thi đấu.
Loại liên kết tham gia hình thành cấu trúc protein:
A. Cấu trúc sơ cấp B. Cấu trúc thứ cấp
B. Cấu trúc bậc ba
D. Cấu trúc siêu trung học E. Cấu tạo.
1. Liên kết hydro giữa các nguyên tử của khung peptide
2. Liên kết yếu giữa các nhóm chức của gốc axit amin
3. Liên kết giữa nhóm α-amino và α-carboxyl của axit amin
8. Chọn những đáp án đúng. Trypsin:
A. Enzym phân giải protein B. Chứa hai miền
B. Thủy phân tinh bột
D. Trang web đang hoạt động nằm giữa các miền. D. Gồm hai chuỗi polypeptide.
9. Chọn những đáp án đúng. Atropin:
A. Chất dẫn truyền thần kinh
B. Cấu trúc tương tự acetylcholine
B. Tương tác với thụ thể H-cholinergic
D. Tăng cường dẫn truyền xung thần kinh thông qua khớp thần kinh cholinergic
D. Chất ức chế cạnh tranh của thụ thể M-cholinergic
10. Chọn những phát biểu đúng. Trong protein:
A. Cấu trúc chính chứa thông tin về cấu trúc của trang hoạt động của nó
B. Trung tâm hoạt động được hình thành ở cấp độ cấu trúc sơ cấp
B. Cấu hình được cố định chắc chắn bằng liên kết cộng hóa trị
D. Vị trí hoạt động có thể tương tác với một nhóm phối tử tương tự
do tính không ổn định về hình dạng của protein D. Những thay đổi trong môi trường có thể ảnh hưởng đến ái lực của hoạt chất
trung tâm phối tử
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
ĐIỀU KHOẢN VÀ KHÁI NIỆM CƠ BẢN
1. Protein, polypeptide, axit amin
2. Cấu trúc protein bậc 1, bậc 2, bậc 3
3. Cấu tạo, cấu hình protein tự nhiên
4. Liên kết cộng hóa trị và liên kết yếu trong protein
5. Khả năng hình dạng
6. Vị trí hoạt động của protein
7. Phối tử
8. Gấp protein
9. Cấu trúc tương tự của phối tử
10. Protein miền
11. Protein đơn giản và phức tạp
12. Chất biến tính protein, chất biến tính
13. Kích hoạt lại protein
Giải quyết vấn đề
“Tổ chức cấu trúc của protein và cơ sở hoạt động của chúng”
1. Chức năng chính của protein - hemoglobin A (HbA) là vận chuyển oxy đến các mô. Trong quần thể người, nhiều dạng protein này được biết đến với các đặc tính và chức năng bị thay đổi - được gọi là huyết sắc tố bất thường. Ví dụ, hemoglobin S, được tìm thấy trong hồng cầu của bệnh nhân mắc bệnh hồng cầu hình liềm (HbS), được phát hiện là có độ hòa tan thấp trong điều kiện áp suất riêng phần của oxy thấp (như trường hợp trong máu tĩnh mạch). Điều này dẫn đến sự hình thành các tập hợp của protein này. Protein mất chức năng, kết tủa và các tế bào hồng cầu có hình dạng không đều (một số có hình liềm) và bị phá hủy nhanh hơn bình thường ở lá lách. Kết quả là bệnh thiếu máu hồng cầu hình liềm phát triển.
Sự khác biệt duy nhất trong cấu trúc bậc một của HbA được tìm thấy ở vùng đầu N của chuỗi β hemoglobin. So sánh các vùng đầu N của sợi β và chỉ ra những thay đổi trong cấu trúc bậc một của protein ảnh hưởng như thế nào đến tính chất và chức năng của nó.
 Đối với điều này:
Đối với điều này:
a) Viết công thức các axit amin phân biệt HbA và so sánh tính chất của các axit amin này (độ phân cực, điện tích).
b) Rút ra kết luận về nguyên nhân làm giảm độ hòa tan và rối loạn vận chuyển oxy vào mô.
2. Hình vẽ cho thấy sơ đồ cấu trúc của protein có trung tâm liên kết với phối tử (trung tâm hoạt động). Giải thích tại sao protein có tính chọn lọc trong việc lựa chọn phối tử. Đối với điều này:
a) nhớ trung tâm hoạt động của protein là gì và xem xét cấu trúc của trung tâm hoạt động của protein như trong hình;
b) viết công thức của các gốc axit amin tạo nên trung tâm hoạt động;
c) vẽ phối tử có thể tương tác đặc hiệu với vị trí hoạt động của protein. Hãy chỉ ra trên đó các nhóm chức có thể tạo liên kết với các gốc axit amin tạo nên trung tâm hoạt động;
d) chỉ ra các loại liên kết phát sinh giữa phối tử và gốc axit amin của trung tâm hoạt động;
e) giải thích tính đặc hiệu của tương tác protein-phối tử dựa trên cơ sở nào.
 3.
Hình vẽ cho thấy vị trí hoạt động của protein và một số phối tử.
3.
Hình vẽ cho thấy vị trí hoạt động của protein và một số phối tử.
Xác định phối tử nào có nhiều khả năng tương tác nhất với vị trí hoạt động của protein và tại sao.
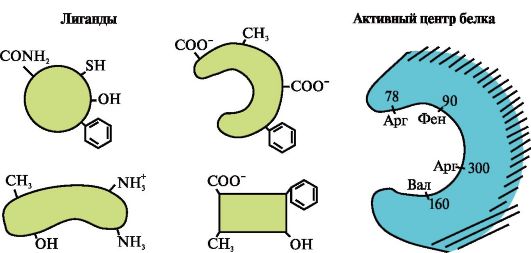 Những loại liên kết nào phát sinh trong quá trình hình thành phức hợp protein-phối tử?
Những loại liên kết nào phát sinh trong quá trình hình thành phức hợp protein-phối tử?
4. Các cấu trúc tương tự của phối tử protein tự nhiên có thể được sử dụng làm thuốc để điều chỉnh hoạt động của protein.
Acetylcholine là chất trung gian dẫn truyền kích thích ở các khớp thần kinh cơ. Khi acetylcholine tương tác với protein - thụ thể của màng sau synap của cơ xương, các kênh ion mở ra và xảy ra hiện tượng co cơ. Ditilin là một loại thuốc được sử dụng trong một số ca phẫu thuật nhằm thư giãn cơ bắp vì nó làm gián đoạn việc truyền xung thần kinh qua các khớp thần kinh cơ. Giải thích cơ chế tác dụng của ditilin như một chất làm giãn cơ. Đối với điều này:
a) viết công thức của acetylcholin và dithilin và so sánh cấu trúc của chúng;
b) mô tả cơ chế tác dụng thư giãn của ditilin.
5. Trong một số bệnh, nhiệt độ cơ thể của người bệnh tăng lên, đây được coi là một phản ứng bảo vệ của cơ thể. Tuy nhiên, nhiệt độ cao có hại cho protein trong cơ thể. Giải thích tại sao ở nhiệt độ trên 40°C, chức năng của protein bị gián đoạn và phát sinh mối đe dọa đối với sự sống con người. Để làm điều này, hãy nhớ:
1) Cấu trúc của protein và các liên kết giữ cấu trúc của nó ở dạng tự nhiên;
2) Cấu trúc và chức năng của protein thay đổi như thế nào khi nhiệt độ tăng?;
3) Cân bằng nội môi là gì và tại sao nó lại quan trọng đối với việc duy trì sức khỏe con người.
Đơn vị mô-đun 2 PROTEIN OLIGOMERICK NHƯ MỤC TIÊU CỦA TÁC ĐỘNG QUY ĐỊNH. ĐA DẠNG CẤU TRÚC VÀ CHỨC NĂNG CỦA PROTEIN. PHƯƠNG PHÁP TÁCH VÀ Tinh chế PROTEIN
Mục tiêu học tập Có khả năng:
1. Sử dụng kiến thức về đặc điểm cấu trúc và chức năng của protein oligomeric để hiểu cơ chế thích nghi điều chỉnh chức năng của chúng.
2. Giải thích vai trò của người đi kèm trong quá trình tổng hợp và duy trì cấu trúc protein trong điều kiện tế bào.
3. Giải thích sự đa dạng của các biểu hiện của sự sống bằng sự đa dạng về cấu trúc và chức năng của các protein được tổng hợp trong cơ thể.
4. Phân tích mối quan hệ giữa cấu trúc của protein và chức năng của chúng bằng cách sử dụng các ví dụ so sánh các hemoprotein liên quan - myoglobin và hemoglobin, cũng như đại diện của năm loại protein thuộc họ immunoglobulin.
5. Vận dụng kiến thức về đặc điểm vật lý, hóa học của protein để lựa chọn phương pháp tinh chế chúng khỏi các protein và tạp chất khác.
6. Giải thích kết quả thành phần định lượng và định tính của protein huyết tương để xác nhận hoặc làm rõ chẩn đoán lâm sàng.
Biết:
1. Đặc điểm cấu trúc của protein oligomeric và cơ chế thích ứng để điều chỉnh chức năng của chúng bằng ví dụ về huyết sắc tố.
2. Cấu trúc và chức năng của người đi kèm và tầm quan trọng của chúng trong việc duy trì cấu trúc tự nhiên của protein trong điều kiện tế bào.
3. Nguyên tắc kết hợp các protein thành các họ dựa trên sự giống nhau về hình dạng và chức năng của chúng bằng ví dụ về globulin miễn dịch.
4. Phương pháp tách protein dựa trên đặc điểm hóa lý của chúng.
5. Điện di huyết tương là phương pháp đánh giá thành phần định tính và định lượng của protein.
CHỦ ĐỀ 1.4. ĐẶC ĐIỂM CẤU TRÚC VÀ CHỨC NĂNG CỦA PROTEIN OLIGOMERIC SỬ DỤNG VÍ DỤ CỦA HEMOGLOBIN
1. Nhiều protein chứa nhiều chuỗi polypeptide. Những protein như vậy được gọi là oligome, và các chuỗi riêng lẻ - các proton. Các proton trong protein oligomeric được kết nối bằng nhiều liên kết không cộng hóa trị yếu (kỵ nước, ion, hydro). Sự tương tác
Promer được thực hiện nhờ sự bổ sung bề mặt tiếp xúc của chúng.
Số lượng proton trong protein oligomeric có thể khác nhau rất nhiều: hemoglobin chứa 4 proton, enzyme aspartate aminotransferase có 12 proton và protein virus khảm thuốc lá chứa 2120 proton được kết nối bằng liên kết không cộng hóa trị. Do đó, protein oligomeric có thể có trọng lượng phân tử rất cao.
Sự tương tác của một proton với các proton khác có thể được coi là trường hợp đặc biệt của tương tác protein-phối tử, vì mỗi proton đóng vai trò là phối tử cho các proton khác. Số lượng và phương pháp liên kết các proton trong protein được gọi là cấu trúc protein bậc bốn.
Protein có thể chứa các proton có cấu trúc giống nhau hoặc khác nhau, ví dụ, homodime là các protein chứa hai proton giống hệt nhau và các dị vòng là các protein chứa hai proton khác nhau.
Nếu protein chứa các proton khác nhau thì các trung tâm liên kết với các phối tử khác nhau có cấu trúc khác nhau có thể được hình thành trên chúng. Khi phối tử liên kết với vị trí hoạt động, chức năng của protein này sẽ được biểu hiện. Một trung tâm nằm trên một proton khác được gọi là allosteric (khác với trung tâm đang hoạt động). Liên hệ phối tử dị lập thể hoặc tác nhân tác động, anh ấy hoàn thành chức năng điều tiết(Hình 1.18). Sự tương tác của trung tâm dị lập thể với cơ quan tác động gây ra những thay đổi về hình dạng trong cấu trúc của toàn bộ protein oligomeric do tính không ổn định về hình dạng của nó. Điều này ảnh hưởng đến ái lực của vị trí hoạt động đối với một phối tử cụ thể và điều chỉnh chức năng của protein đó. Sự thay đổi về hình dạng và chức năng của tất cả các proton trong quá trình tương tác giữa protein oligomeric với ít nhất một phối tử được gọi là những thay đổi về hình dạng hợp tác. Các tác nhân làm tăng cường chức năng của protein được gọi là kích hoạt, và các tác nhân ức chế chức năng của nó - chất ức chế.
Do đó, protein oligomeric, cũng như các protein có cấu trúc miền, có một đặc tính mới so với protein đơn phân - khả năng điều chỉnh các chức năng phân lập (điều chỉnh bằng cách gắn các phối tử khác nhau vào protein). Điều này có thể được nhận thấy bằng cách so sánh cấu trúc và chức năng của hai loại protein phức tạp có liên quan chặt chẽ với nhau là myoglobin và hemoglobin.
 Cơm. 1.18. Sơ đồ cấu trúc của protein dimeric
Cơm. 1.18. Sơ đồ cấu trúc của protein dimeric
2. Sự hình thành cấu trúc không gian và chức năng của myoglobin.
Myoglobin (Mb) là một loại protein có trong cơ đỏ, chức năng chính của nó là tạo ra nguồn dự trữ O 2 cần thiết cho hoạt động cơ bắp cường độ cao. Mb là một protein phức tạp chứa một phần protein - apoMb và một phần không phải protein - heme. Cấu trúc bậc một của apoMB xác định cấu trúc hình cầu nhỏ gọn của nó và cấu trúc của trung tâm hoạt động, nơi gắn phần phi protein của myoglobin, heme. Oxy từ máu đến cơ liên kết với heme Fe+2 trong myoglobin. Mb là một protein đơn phân có ái lực rất cao với O 2, do đó sự giải phóng oxy bởi myoglobin chỉ xảy ra khi cơ bắp hoạt động mạnh, khi áp suất riêng phần của O 2 giảm mạnh.
Sự hình thành cấu hình Mv.Ở cơ đỏ, trên ribosome trong quá trình dịch mã, cấu trúc bậc một của MB được tổng hợp, thể hiện bằng một trình tự cụ thể gồm 153 gốc axit amin. Cấu trúc thứ cấp của Mb chứa tám chuỗi xoắn ốc, được gọi bằng chữ cái Latinh từ A đến H, giữa chúng có các vùng không xoắn ốc. Cấu trúc bậc ba của Mb có dạng một cầu nhỏ gọn, trong phần lõm của nó có trung tâm hoạt động nằm giữa các chuỗi xoắn ốc F và E (Hình 1.19).
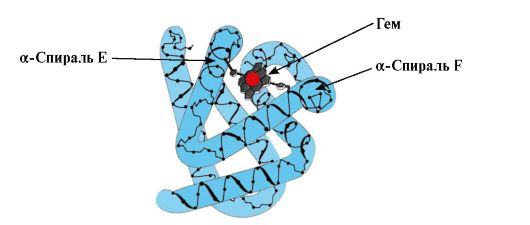 Cơm. 1.19. Cấu trúc myoglobin
Cơm. 1.19. Cấu trúc myoglobin
3. Đặc điểm cấu tạo và hoạt động của trung tâm hoạt động MV. Trung tâm hoạt động của Mb được hình thành chủ yếu bởi các gốc axit amin kỵ nước, cách xa nhau trong cấu trúc bậc một (ví dụ, Tri 3 9 và Fen 138) Ít tan trong các phối tử nước - heme và O 2 - gắn vào trung tâm hoạt động. Heme là một phối tử đặc hiệu của apoMB (Hình 1.20), cơ sở của nó được tạo thành từ bốn vòng pyrrole được nối với nhau bằng cầu methenyl; ở trung tâm có một nguyên tử Fe+2 được nối với các nguyên tử nitơ của vòng pyrrole bằng bốn liên kết phối trí. Ở trung tâm hoạt động của Mb, ngoài các gốc axit amin kỵ nước còn có dư lượng của hai axit amin có gốc ưa nước - Gis E 7(Gis 64) và GIS F 8(của anh ấy là 93) (Hình 1.21).
 Cơm. 1,20. Cấu trúc của heme - phần phi protein của myoglobin và hemoglobin
Cơm. 1,20. Cấu trúc của heme - phần phi protein của myoglobin và hemoglobin
 Cơm. 1,21. Vị trí của heme và O2 trong trung tâm hoạt động của các proton apomyoglobin và hemoglobin
Cơm. 1,21. Vị trí của heme và O2 trong trung tâm hoạt động của các proton apomyoglobin và hemoglobin
Heme liên kết cộng hóa trị với F8 của anh ấy thông qua một nguyên tử sắt. O 2 gắn vào thanh sắt ở phía bên kia của mặt phẳng heme. E 7 của anh ấy cần thiết cho sự định hướng chính xác của O 2 và tạo điều kiện thuận lợi cho việc bổ sung oxy vào heme Fe + 2
GIS F 8 tạo thành liên kết phối trí với Fe+2 và cố định chắc chắn heme ở trung tâm hoạt động. Gis E 7 cần thiết cho sự định hướng chính xác ở trung tâm hoạt động của phối tử khác - O 2 trong quá trình tương tác với heme Fe + 2. Môi trường vi mô của heme tạo điều kiện cho sự liên kết mạnh mẽ nhưng thuận nghịch của O 2 với Fe + 2 và ngăn nước xâm nhập vào vị trí hoạt động kỵ nước, có thể dẫn đến quá trình oxy hóa nó thành Fe + 3.
Cấu trúc đơn phân của Mb và trung tâm hoạt động của nó quyết định ái lực cao của protein đối với O 2.
4. Cấu trúc oligomeric của Hb và sự điều hòa ái lực của Hb với phối tử O 2. Huyết sắc tố người- một họ protein, như myoglobin, liên quan đến các protein phức tạp (hemoprotein). Chúng có cấu trúc tứ giác và chứa hai chuỗi α, nhưng khác nhau về cấu trúc của hai chuỗi polypeptide còn lại (chuỗi 2α-, 2x). Cấu trúc của chuỗi polypeptide thứ hai xác định đặc điểm hoạt động của các dạng Hb này. Khoảng 98% lượng huyết sắc tố trong hồng cầu của người trưởng thành là huyết sắc tố A(chuỗi 2α-, 2p).
Trong quá trình phát triển của thai nhi, hai loại huyết sắc tố chính có chức năng: Hb phôi thai(2α, 2ε), được tìm thấy trong giai đoạn đầu phát triển của thai nhi, và huyết sắc tố F (thai nhi)- (2α, 2γ), thay thế huyết sắc tố sớm của thai nhi vào tháng thứ sáu của quá trình phát triển trong tử cung và chỉ sau khi sinh mới được thay thế bằng Hb A.
HB A là một protein liên quan đến myoglobin (MB) được tìm thấy trong tế bào hồng cầu ở người trưởng thành. Cấu trúc của các proton riêng lẻ của nó tương tự như cấu trúc của myoglobin. Cấu trúc bậc hai và bậc ba của các proton myoglobin và hemoglobin rất giống nhau, mặc dù thực tế là trong cấu trúc bậc một của chuỗi polypeptide của chúng chỉ có 24 gốc axit amin giống hệt nhau (cấu trúc bậc hai của các proton myoglobin, như myoglobin, chứa tám chuỗi xoắn ốc, được ký hiệu bằng các chữ cái Latinh từ A đến H và cấu trúc bậc ba có dạng hình cầu nhỏ gọn). Nhưng không giống như myoglobin, hemoglobin có cấu trúc oligomeric, bao gồm bốn chuỗi polypeptide được nối với nhau bằng liên kết không cộng hóa trị (Hình 1.22).
Mỗi proton Hb được liên kết với một phần không phải protein - heme và các proton lân cận. Sự kết nối giữa phần protein của Hb với heme tương tự như sự kết nối của myoglobin: ở trung tâm hoạt động của protein, phần kỵ nước của heme được bao quanh bởi các gốc axit amin kỵ nước ngoại trừ His F 8 và His E 7, nằm ở cả hai phía của mặt phẳng heme và đóng vai trò tương tự trong hoạt động của protein và liên kết của nó với oxy (xem cấu trúc của myoglobin).
 Cơm. 1,22. Cấu trúc oligomeric của hemoglobin
Cơm. 1,22. Cấu trúc oligomeric của hemoglobin
Bên cạnh đó, Gis E 7 thực hiện một chức năng quan trọng vai trò bổ sung trong hoạt động của Nv. Heme tự do có ái lực với CO cao hơn 25.000 lần so với O2. CO được hình thành với số lượng nhỏ trong cơ thể và do có ái lực cao với heme, nó có thể làm gián đoạn quá trình vận chuyển O 2 cần thiết cho sự sống của tế bào. Tuy nhiên, trong thành phần của hemoglobin, ái lực của heme với carbon monoxide chỉ vượt quá ái lực với O 2 200 lần do sự hiện diện của His E 7 ở trung tâm hoạt động. Phần axit amin còn lại tạo điều kiện tối ưu cho sự liên kết của heme với O 2 và làm suy yếu sự tương tác của heme với CO.
5. Chức năng chính của HB là vận chuyển O2 từ phổi đến các mô. Không giống như myoglobin đơn phân, có ái lực rất cao với O2 và thực hiện chức năng lưu trữ oxy trong cơ đỏ, cấu trúc oligomeric của hemoglobin cung cấp:
1) bão hòa nhanh chóng HB với oxy trong phổi;
2) khả năng HB giải phóng oxy vào các mô ở áp suất riêng phần O 2 tương đối cao (20-40 mm Hg);
3) khả năng điều chỉnh ái lực của Hb với O 2.
6. Những thay đổi hợp tác về hình dạng của các proton proton trong huyết sắc tố làm tăng tốc độ liên kết của O 2 trong phổi và giải phóng nó vào các mô. Trong phổi, áp suất riêng phần cao của O 2 thúc đẩy sự liên kết của nó với Hb ở vị trí hoạt động của bốn proton (2α và 2β). Trung tâm hoạt động của mỗi proton, như trong myoglobin, nằm giữa hai chuỗi xoắn α (F và E) trong một túi kỵ nước. Nó chứa một phần không phải protein - heme, được gắn vào phần protein bằng nhiều tương tác kỵ nước yếu và một liên kết bền giữa Fe 2 + heme và His F 8 (xem hình 1.21).
Trong deoxyhemoglobin, do liên kết với His F 8 nên nguyên tử Fe 2 + nhô ra khỏi mặt phẳng heme về phía histidine. Sự liên kết của O 2 với Fe 2 + xảy ra ở phía bên kia của heme trong vùng His E 7 bằng một liên kết phối hợp tự do duy nhất. E 7 của ông cung cấp các điều kiện tối ưu cho sự liên kết của O 2 với sắt heme.
Việc bổ sung O 2 vào nguyên tử Fe + 2 của một proton sẽ khiến nó di chuyển vào mặt phẳng heme, sau đó là dư lượng histidine liên kết với nó
 Cơm. 1,23. Thay đổi cấu hình của protone hemoglobin khi kết hợp với O 2
Cơm. 1,23. Thay đổi cấu hình của protone hemoglobin khi kết hợp với O 2
Điều này dẫn đến sự thay đổi về hình dạng của tất cả các chuỗi polypeptide do tính không ổn định về hình dạng của chúng. Việc thay đổi hình dạng của các chuỗi khác tạo điều kiện thuận lợi cho sự tương tác của chúng với các phân tử O 2 tiếp theo.
Phân tử O 2 thứ tư gắn vào hemoglobin dễ dàng hơn 300 lần so với phân tử đầu tiên (Hình 1.24).
 Cơm. 1,24. Những thay đổi hợp tác về hình dạng của các proton proton trong quá trình tương tác với O2
Cơm. 1,24. Những thay đổi hợp tác về hình dạng của các proton proton trong quá trình tương tác với O2
Trong các mô, mỗi phân tử O 2 tiếp theo bị tách ra dễ dàng hơn phân tử trước đó, cũng là do sự thay đổi hợp tác về hình dạng của các proton.
7. CO 2 và H+ được hình thành trong quá trình dị hóa các chất hữu cơ, làm giảm ái lực của hemoglobin đối với O 2 tương ứng với nồng độ của chúng. Năng lượng cần thiết cho hoạt động của tế bào được tạo ra chủ yếu ở ty thể trong quá trình oxy hóa các chất hữu cơ sử dụng O 2 được vận chuyển từ phổi bởi huyết sắc tố. Kết quả của quá trình oxy hóa các chất hữu cơ, sản phẩm cuối cùng của quá trình phân hủy chúng được hình thành: CO 2 và K 2 O, lượng này tỷ lệ thuận với cường độ của quá trình oxy hóa đang diễn ra.
CO 2 khuếch tán từ tế bào vào máu và xâm nhập vào hồng cầu, tại đây, dưới tác dụng của enzyme carbanhydrase, nó được chuyển thành axit carbonic. Axit yếu này phân ly thành proton và ion bicarbonate.
H+ có khả năng kết hợp các gốc tự do của His 14 6 trong chuỗi α- và β của hemoglobin, tức là ở những khu vực xa heme. Sự proton hóa của hemoglobin làm giảm ái lực của nó với O 2, thúc đẩy quá trình loại bỏ O 2 khỏi oxyHb, hình thành deoxyHb và tăng cung cấp oxy cho các mô tỷ lệ với số lượng proton được hình thành (Hình 1.25).
Sự gia tăng lượng oxy giải phóng tùy thuộc vào sự gia tăng nồng độ H+ trong hồng cầu được gọi là hiệu ứng Bohr (được đặt theo tên của nhà sinh lý học người Đan Mạch Christian Bohr, người đầu tiên phát hiện ra hiệu ứng này).
Trong phổi, áp suất riêng phần cao của oxy thúc đẩy sự liên kết của nó với deoxyHb, làm giảm ái lực của protein với H+. Proton được giải phóng dưới tác dụng của axit cacbonic phản ứng với bicarbonat tạo thành CO 2 và H 2 O

 Cơm. 1,25. Sự phụ thuộc ái lực của Hb với O 2 vào nồng độ CO 2 và proton (hiệu ứng Bohr):
Cơm. 1,25. Sự phụ thuộc ái lực của Hb với O 2 vào nồng độ CO 2 và proton (hiệu ứng Bohr):
MỘT- ảnh hưởng của nồng độ CO 2 và H+ đến việc giải phóng O 2 khỏi phức hợp HB (hiệu ứng Bohr); B- oxy hóa deoxyhemoglobin trong phổi, hình thành và giải phóng CO 2.
CO 2 thu được đi vào khoang phế nang và được loại bỏ bằng không khí thở ra. Do đó, lượng oxy do hemoglobin giải phóng trong các mô được điều hòa bởi các sản phẩm dị hóa của các chất hữu cơ: sự phân hủy các chất càng mạnh, ví dụ như khi tập thể dục, nồng độ CO 2 và H + càng cao và càng nhiều oxy các mô nhận được do giảm ái lực của Hb với O 2.
8. Điều hòa dị lập thể ái lực của Hb với O2 bằng phối tử - 2,3-bisphosphoglycerate. Trong hồng cầu, phối tử dị lập thể của hemoglobin là 2,3-bisphosphoglycerate (2,3-BPG), được tổng hợp từ sản phẩm của quá trình oxy hóa glucose - 1,3-bisphosphoglycerate. Trong điều kiện bình thường, nồng độ 2,3-BPG cao và có thể sánh ngang với nồng độ Hb. 2,3-BPG có điện tích âm mạnh -5.
 Bisphosphoglycerate trong mao mạch mô, liên kết với deoxyhemoglobin, làm tăng giải phóng oxy vào mô, làm giảm ái lực của Hb với O 2.
Bisphosphoglycerate trong mao mạch mô, liên kết với deoxyhemoglobin, làm tăng giải phóng oxy vào mô, làm giảm ái lực của Hb với O 2.
Ở trung tâm của phân tử tetrameric hemoglobin là một khoang. Nó được hình thành bởi dư lượng axit amin của cả bốn proton (xem hình 1.22). Trong mao mạch mô, sự proton hóa của Hb (hiệu ứng Bohr) dẫn đến sự đứt gãy liên kết giữa sắt heme và O2. Trong một phân tử
deoxyhemoglobin, so với oxyhemoglobin, xuất hiện các liên kết ion bổ sung kết nối các proton, do đó kích thước của khoang trung tâm tăng lên so với oxyhemoglobin. Khoang trung tâm là nơi gắn 2,3-BPG với hemoglobin. Do sự khác biệt về kích thước của khoang trung tâm nên 2,3-BPG chỉ có thể gắn vào deoxyhemoglobin.
2,3-BPG tương tác với hemoglobin ở vị trí cách xa trung tâm hoạt động của protein và thuộc về phân lập thể phối tử (điều hòa), và khoang trung tâm của Hb là trung tâm dị lập thể. 2,3-BPG có điện tích âm mạnh và tương tác với 5 nhóm tích điện dương của hai chuỗi β của Hb: nhóm α-amino đầu N của Val và gốc Lys 82 His 143 (Hình 1.26).
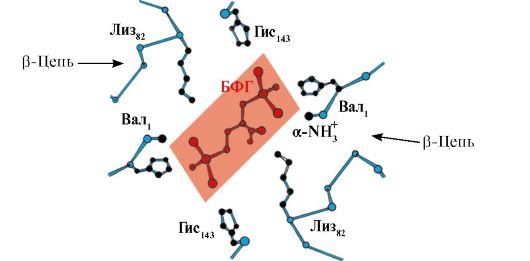 Cơm. 1.26. BPG trong khoang trung tâm của deoxyhemoglobin
Cơm. 1.26. BPG trong khoang trung tâm của deoxyhemoglobin
BPG liên kết với ba nhóm tích điện dương trên mỗi sợi β.
Trong các mao mạch mô, deoxyhemoglobin thu được tương tác với 2,3-BPG và liên kết ion được hình thành giữa các gốc tích điện dương của chuỗi β và phối tử tích điện âm, làm thay đổi cấu trúc của protein và làm giảm ái lực của Hb với O2. . Sự giảm ái lực của Hb với O 2 góp phần giải phóng O 2 vào mô hiệu quả hơn.
Trong phổi, ở áp suất riêng phần cao, oxy tương tác với Hb, kết hợp với sắt heme; trong trường hợp này, cấu trúc của protein thay đổi, khoang trung tâm giảm đi và 2,3-BPG bị dịch chuyển khỏi trung tâm dị lập thể
Do đó, protein oligomeric có những đặc tính mới so với protein đơn phân. Gắn các phối tử tại các vị trí
cách xa nhau về mặt không gian (alosteric), có thể gây ra những thay đổi về hình dạng trong toàn bộ phân tử protein. Do sự tương tác với các phối tử điều hòa, xảy ra sự thay đổi về hình dạng và sự thích ứng chức năng của phân tử protein với những thay đổi của môi trường.
CHỦ ĐỀ 1.5. DUY TRÌ CẤU HÌNH TỰ NHIÊN CỦA PROTEIN TRONG ĐIỀU KIỆN TẾ BÀO
Trong tế bào, trong quá trình tổng hợp chuỗi polypeptide, sự vận chuyển chúng qua màng đến các phần tương ứng của tế bào, trong quá trình gấp nếp (hình thành cấu trúc tự nhiên) và trong quá trình lắp ráp các protein oligomeric, cũng như trong quá trình hoạt động, trung gian của chúng. , sự hình thành dễ kết tụ, không ổn định phát sinh trong cấu trúc protein. Các gốc kỵ nước, thường ẩn bên trong phân tử protein ở dạng tự nhiên, xuất hiện trên bề mặt ở dạng không ổn định và có xu hướng kết hợp với các nhóm protein khác hòa tan kém trong nước. Trong tế bào của tất cả các sinh vật đã biết, người ta đã tìm thấy các protein đặc biệt đảm bảo sự gấp cuộn tối ưu của protein tế bào, ổn định cấu trúc tự nhiên của chúng trong quá trình hoạt động và quan trọng nhất là duy trì cấu trúc và chức năng của protein nội bào khi cân bằng nội môi bị xáo trộn. Những protein này được gọi là "người đi kèm" có nghĩa là "vú em" trong tiếng Pháp.
1. Người đi kèm phân tử và vai trò của chúng trong việc ngăn chặn sự biến tính protein.
Người đi kèm (CH) được phân loại theo khối lượng của các tiểu đơn vị của họ. Người đi kèm có trọng lượng phân tử cao có khối lượng từ 60 đến 110 kDa. Trong đó, 3 lớp được nghiên cứu nhiều nhất là Sh-60, Sh-70 và Sh-90. Mỗi lớp bao gồm một họ protein có liên quan. Do đó, Sh-70 bao gồm các protein có trọng lượng phân tử từ 66 đến 78 kDa. Người đi kèm trọng lượng phân tử thấp có trọng lượng phân tử từ 40 đến 15 kDa.
Trong số những người đi kèm có cấu thành protein, mức tổng hợp cơ bản cao không phụ thuộc vào tác động căng thẳng lên tế bào của cơ thể và cảm ứng, sự tổng hợp trong điều kiện bình thường là yếu, nhưng tăng mạnh khi bị căng thẳng. Những người đi kèm cảm ứng còn được gọi là “protein sốc nhiệt” vì chúng lần đầu tiên được phát hiện trong các tế bào tiếp xúc với nhiệt độ cao. Trong tế bào, do nồng độ protein cao nên việc tái kích hoạt tự phát các protein bị biến tính một phần là rất khó khăn. Sh-70 có thể ngăn chặn sự biến tính và giúp khôi phục cấu trúc tự nhiên của protein. Người đi kèm phân tử-70- một loại protein có tính bảo tồn cao được tìm thấy trong tất cả các phần của tế bào: tế bào chất, nhân, mạng lưới nội chất, ty thể. Ở đầu carboxyl của chuỗi polypeptide đơn Ш-70 có một vùng là rãnh có khả năng tương tác với các peptide có chiều dài
từ 7 đến 9 gốc axit amin được làm giàu bằng gốc kỵ nước. Những vùng như vậy trong protein hình cầu xuất hiện khoảng 16 axit amin. Sh-70 có khả năng bảo vệ protein khỏi sự bất hoạt của nhiệt độ và khôi phục cấu trúc cũng như hoạt động của protein bị biến tính một phần.
2. Vai trò của người đi kèm trong việc gấp protein. Trong quá trình tổng hợp protein trên ribosome, vùng đầu N của polypeptide được tổng hợp trước vùng đầu C. Để hình thành cấu trúc tự nhiên, cần có trình tự axit amin hoàn chỉnh của protein. Trong quá trình tổng hợp protein, chaperones-70, do cấu trúc của trung tâm hoạt động của chúng, có khả năng đóng các vùng polypeptide dễ bị kết tụ, được làm giàu bằng các gốc axit amin kỵ nước cho đến khi quá trình tổng hợp hoàn tất (Hình 1.27, A). ).
 Cơm. 1.27. Sự tham gia của người đi kèm trong quá trình gấp protein
Cơm. 1.27. Sự tham gia của người đi kèm trong quá trình gấp protein
A - sự tham gia của người đi kèm-70 trong việc ngăn chặn các tương tác kỵ nước giữa các phần của polypeptide tổng hợp; B - sự hình thành cấu trúc tự nhiên của protein trong phức hợp chaperone
Nhiều protein phân tử cao có cấu trúc phức tạp, chẳng hạn như cấu trúc miền, gấp lại trong một không gian đặc biệt được hình thành bởi Sh-60. Ш-60 hoạt động như một phức hợp oligomeric bao gồm 14 tiểu đơn vị. Chúng tạo thành hai vòng rỗng, mỗi vòng bao gồm bảy tiểu đơn vị, các vòng này được kết nối với nhau. Mỗi tiểu đơn vị Sh-60 bao gồm ba miền: đỉnh (đỉnh), được làm giàu bằng các gốc kỵ nước đối diện với khoang của vòng, trung gian và xích đạo (Hình 1.28).
 Cơm. 1,28. Cấu trúc của phức hợp chaperonin gồm 14 Ш-60
Cơm. 1,28. Cấu trúc của phức hợp chaperonin gồm 14 Ш-60
A - nhìn từ bên; B - nhìn từ trên xuống
Các protein tổng hợp, có các thành phần trên bề mặt đặc trưng của các phân tử chưa được gấp nếp, đặc biệt là các gốc kỵ nước, đi vào khoang của các vòng chaperone. Trong môi trường cụ thể của các khoang này, các cấu hình có thể được tìm kiếm cho đến khi tìm thấy cấu hình duy nhất có lợi nhất về mặt năng lượng (Hình 1.27, B). Sự hình thành cấu hình và giải phóng protein đi kèm với quá trình thủy phân ATP ở vùng xích đạo. Thông thường, việc gấp nếp phụ thuộc vào người đi kèm như vậy đòi hỏi một lượng năng lượng đáng kể.
Ngoài việc tham gia vào việc hình thành cấu trúc ba chiều của protein và tái tạo protein bị biến tính một phần, người đi kèm cũng cần thiết cho sự xuất hiện của các quá trình cơ bản như lắp ráp protein oligomeric, nhận biết và vận chuyển protein bị biến tính vào lysosome, vận chuyển protein qua màng và tham gia điều hòa hoạt động của phức hợp protein.
CHỦ ĐỀ 1.6. CÁC LOẠI PROTEIN. CÁC HỌ PROTEIN: VÍ DỤ VỀ IMUNOGLOBULIN
1. Protein đóng vai trò quyết định trong đời sống của từng tế bào và toàn bộ cơ thể đa bào, và chức năng của chúng rất đa dạng. Điều này được xác định bởi các đặc điểm của cấu trúc bậc một và sự phù hợp của protein, cấu trúc độc đáo của trung tâm hoạt động và khả năng liên kết các phối tử cụ thể.
Chỉ một phần rất nhỏ trong số tất cả các biến thể có thể có của chuỗi peptide có thể áp dụng cấu trúc không gian ổn định; số đông
trong số chúng có thể có nhiều hình dạng với năng lượng Gibbs xấp xỉ nhau, nhưng với tính chất khác nhau. Cấu trúc bậc một của hầu hết các protein được biết đến được lựa chọn bởi quá trình tiến hóa sinh học đảm bảo tính ổn định đặc biệt của một trong những cấu hình đó, xác định các đặc điểm hoạt động của protein này.
2. Họ protein. Trong cùng một loài sinh vật, việc thay thế các gốc axit amin có thể dẫn đến sự xuất hiện của các protein khác nhau thực hiện các chức năng liên quan và có trình tự axit amin tương đồng. Các protein liên quan như vậy có cấu hình tương tự nhau một cách đáng kinh ngạc: số lượng và vị trí tương đối của các cấu trúc xoắn α và/hoặc cấu trúc β, và hầu hết các vòng xoắn và độ uốn của chuỗi polypeptide đều tương tự hoặc giống hệt nhau. Các protein có các vùng tương đồng của chuỗi polypeptide, có cấu trúc tương tự và các chức năng liên quan được phân loại vào các họ protein. Ví dụ về họ protein: serine proteinase, họ immunoglobulin, họ myoglobin.
Proteinase huyết thanh- một họ protein thực hiện chức năng của các enzyme phân giải protein. Chúng bao gồm các enzyme tiêu hóa - chymotrypsin, trypsin, elastase và nhiều yếu tố đông máu. Những protein này có các axit amin giống nhau ở 40% vị trí và có cấu trúc rất giống nhau (Hình 1.29).

Cơm. 1,29. Cấu trúc không gian của elastase (A) và chymotrypsin (B)
Một số thay thế axit amin đã dẫn đến những thay đổi về tính đặc hiệu cơ chất của các protein này và sự xuất hiện của sự đa dạng về chức năng trong họ.
3. Họ globulin miễn dịch. Trong hoạt động của hệ thống miễn dịch, các protein thuộc siêu họ globulin miễn dịch đóng một vai trò rất lớn, bao gồm ba họ protein:
Kháng thể (globulin miễn dịch);
Thụ thể tế bào lympho T;
Protein của phức hợp tương hợp mô chính - MHC lớp 1 và 2 (Phức hợp tương hợp mô chính).
Tất cả các protein này đều có cấu trúc miền, bao gồm các miền giống miễn dịch tương đồng và thực hiện các chức năng tương tự: chúng tương tác với các cấu trúc lạ, hòa tan trong máu, bạch huyết hoặc dịch gian bào (kháng thể), hoặc nằm trên bề mặt tế bào (riêng hoặc nước ngoài).
4. Kháng thể- các protein đặc hiệu được tạo ra bởi tế bào lympho B để đáp ứng với sự xâm nhập của cấu trúc lạ vào cơ thể, được gọi là kháng nguyên.
Đặc điểm cấu trúc của kháng thể
Các phân tử kháng thể đơn giản nhất bao gồm bốn chuỗi polypeptide: hai chuỗi nhẹ giống hệt nhau - L, chứa khoảng 220 axit amin và hai chuỗi nặng giống hệt nhau - H, gồm 440-700 axit amin. Tất cả bốn chuỗi trong phân tử kháng thể được nối với nhau bằng nhiều liên kết không cộng hóa trị và bốn liên kết disulphide (Hình 1.30).
Chuỗi nhẹ kháng thể bao gồm hai miền: miền biến đổi (VL), nằm ở vùng đầu N của chuỗi polypeptide, và miền cố định (CL), nằm ở đầu C. Chuỗi nặng thường có bốn vùng: một vùng biến đổi (VH), nằm ở đầu N và ba vùng cố định (CH1, CH2, CH3) (xem Hình 1.30). Mỗi miền globulin miễn dịch có cấu trúc thượng tầng tấm β trong đó hai phần cystein được liên kết bằng liên kết disulfide.
Giữa hai miền cố định CH1 và CH2 có một vùng chứa số lượng lớn gốc proline, ngăn cản sự hình thành cấu trúc bậc hai và sự tương tác của các chuỗi H lân cận trong đoạn này. Vùng bản lề này mang lại sự linh hoạt cho phân tử kháng thể. Giữa các miền biến đổi của chuỗi nặng và chuỗi nhẹ có hai vị trí liên kết kháng nguyên giống hệt nhau (vị trí hoạt động để liên kết kháng nguyên), do đó các kháng thể như vậy thường được gọi là hóa trị hai. Không phải toàn bộ trình tự axit amin của các vùng biến đổi của cả hai chuỗi đều tham gia vào quá trình liên kết kháng nguyên với kháng thể mà chỉ có 20-30 axit amin nằm trong vùng siêu biến của mỗi chuỗi. Chính những vùng này quyết định khả năng duy nhất của từng loại kháng thể trong việc tương tác với kháng nguyên bổ sung tương ứng.
Kháng thể là một trong những tuyến phòng thủ của cơ thể chống lại sự xâm nhập của các sinh vật lạ. Chức năng của chúng có thể được chia thành hai giai đoạn: giai đoạn đầu tiên là nhận biết và liên kết kháng nguyên trên bề mặt của sinh vật lạ, điều này có thể thực hiện được nhờ sự hiện diện của các vị trí liên kết kháng nguyên trong cấu trúc kháng thể; giai đoạn thứ hai là bắt đầu quá trình bất hoạt và tiêu diệt kháng nguyên. Độ đặc hiệu của giai đoạn thứ hai phụ thuộc vào loại kháng thể. Có năm loại chuỗi nặng, khác nhau về cấu trúc của các miền không đổi: α, δ, ε, γ và μ, theo đó phân biệt năm loại globulin miễn dịch: A, D, E, G và M.
Đặc điểm cấu trúc của chuỗi nặng mang lại cho vùng bản lề và vùng đầu C của chuỗi nặng đặc điểm cấu tạo của từng loại. Sau khi kháng nguyên liên kết với kháng thể, những thay đổi về hình dạng trong các miền cố định sẽ xác định con đường loại bỏ kháng nguyên.
 Cơm. 1. 30. Cấu trúc miền của IgG
Cơm. 1. 30. Cấu trúc miền của IgG
Globulin miễn dịch M
Globulin miễn dịch M có hai dạng.
Dạng đơn phân- Loại kháng thể thứ nhất được tạo ra bởi tế bào lympho B đang phát triển. Sau đó, nhiều tế bào B chuyển sang sản xuất các loại kháng thể khác, nhưng có cùng vị trí gắn kháng nguyên. IgM được nhúng vào màng và hoạt động như một thụ thể nhận biết kháng nguyên. Sự tích hợp của IgM vào màng tế bào có thể xảy ra do sự hiện diện của 25 gốc axit amin kỵ nước ở phần đuôi của vùng.
Dạng bài tiết của IgM chứa năm tiểu đơn vị đơn phân liên kết với nhau bằng liên kết disulfide và chuỗi polypeptide J bổ sung (Hình 1.31). Các chuỗi monome nặng ở dạng này không chứa đuôi kỵ nước. Pentamer có 10 vị trí gắn kháng nguyên và do đó có hiệu quả trong việc nhận biết và loại bỏ kháng nguyên lần đầu tiên xâm nhập vào cơ thể. Dạng bài tiết của IgM là loại kháng thể chính được tiết vào máu trong phản ứng miễn dịch cơ bản. Sự gắn kết của IgM với kháng nguyên làm thay đổi cấu trúc của IgM và tạo ra sự liên kết của nó với thành phần protein đầu tiên của hệ thống bổ thể (hệ thống bổ sung là một tập hợp các protein liên quan đến việc phá hủy kháng nguyên) và kích hoạt hệ thống này. Nếu kháng nguyên nằm trên bề mặt của vi sinh vật, hệ thống bổ thể sẽ gây ra sự phá vỡ tính toàn vẹn của màng tế bào và làm chết tế bào vi khuẩn.
Globulin miễn dịch G
Về mặt định lượng, loại globulin miễn dịch này chiếm ưu thế trong máu (75% tổng số Igs). IgG - monome, loại kháng thể chính được tiết vào máu trong phản ứng miễn dịch thứ cấp. Sau sự tương tác của IgG với kháng nguyên bề mặt của vi sinh vật, phức hợp kháng nguyên-kháng thể có khả năng liên kết và kích hoạt các protein của hệ thống bổ sung hoặc có thể tương tác với các thụ thể cụ thể của đại thực bào và bạch cầu trung tính. Tương tác với các tế bào dẫn thực bào
 Cơm. 1.31. Cấu trúc dạng bài tiết của IgM
Cơm. 1.31. Cấu trúc dạng bài tiết của IgM
đến sự hấp thụ phức hợp kháng nguyên-kháng thể và sự phá hủy chúng trong các phagosome của tế bào. IgG là loại kháng thể duy nhất có khả năng xuyên qua hàng rào nhau thai và bảo vệ thai nhi khỏi bị nhiễm trùng trong tử cung.
Globulin miễn dịch A
Nhóm kháng thể chính có trong dịch tiết (sữa, nước bọt, dịch tiết của đường hô hấp và đường ruột). IgA được tiết ra chủ yếu ở dạng dimeric, trong đó các monome được liên kết với nhau thông qua chuỗi J bổ sung (Hình 1.32).
IgA không tương tác với hệ thống bổ sung và tế bào thực bào, nhưng bằng cách liên kết với vi sinh vật, kháng thể ngăn chặn sự gắn kết của chúng với tế bào biểu mô và xâm nhập vào cơ thể.
Globulin miễn dịch E
Globulin miễn dịch E được đại diện bởi các monome trong đó chuỗi ε nặng chứa, giống như chuỗi μ của globulin miễn dịch M, một vùng biến đổi và bốn miền cố định. Sau khi bài tiết, IgE liên kết với
 Cơm. 1,32. Cấu trúc của IgA
Cơm. 1,32. Cấu trúc của IgA
Vùng đầu C có các thụ thể tương ứng trên bề mặt tế bào mast và bạch cầu ái kiềm. Kết quả là chúng trở thành thụ thể kháng nguyên trên bề mặt tế bào này (Hình 1.33).
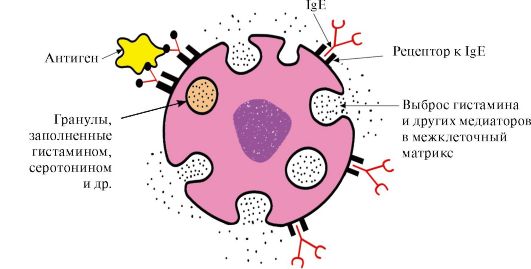 Cơm. 1,33. Tương tác của IgE với kháng nguyên trên bề mặt tế bào mast
Cơm. 1,33. Tương tác của IgE với kháng nguyên trên bề mặt tế bào mast
Sau khi kháng nguyên gắn vào vị trí gắn kháng nguyên tương ứng của IgE, các tế bào sẽ nhận được tín hiệu tiết ra các hoạt chất sinh học (histamine, serotonin), những chất này chịu trách nhiệm chính cho sự phát triển của phản ứng viêm và biểu hiện các phản ứng dị ứng như: hen suyễn, nổi mề đay, sốt cỏ khô.
Globulin miễn dịch D
Globulin miễn dịch D được tìm thấy với số lượng rất nhỏ trong huyết thanh; chúng là các monome. Chuỗi δ nặng có một miền biến đổi và ba miền cố định. IgD hoạt động như các thụ thể của tế bào lympho B; các chức năng khác vẫn chưa được biết rõ. Sự tương tác của các kháng nguyên cụ thể với các thụ thể trên bề mặt tế bào lympho B (IgD) dẫn đến việc truyền các tín hiệu này vào tế bào và kích hoạt các cơ chế đảm bảo sự tăng sinh của một dòng tế bào lympho nhất định.
CHỦ ĐỀ 1.7. ĐẶC TÍNH VẬT LÝ VÀ HÓA HỌC CỦA PROTEIN VÀ PHƯƠNG PHÁP TÁCH chúng
1. Các protein riêng lẻ khác nhau về tính chất vật lý và hóa học:
Hình dạng của phân tử;
Trọng lượng phân tử;
Tổng điện tích, độ lớn của nó phụ thuộc vào tỷ lệ giữa các nhóm axit amin anion và cation;
Tỷ lệ các gốc axit amin phân cực và không phân cực trên bề mặt phân tử;
Mức độ kháng lại các tác nhân biến tính khác nhau.
2. Độ hòa tan của protein phụ thuộc vào các đặc tính của protein được liệt kê ở trên, cũng như thành phần của môi trường hòa tan protein (giá trị pH, thành phần muối, nhiệt độ, sự có mặt của các chất hữu cơ khác có thể tương tác với protein). Lượng điện tích của phân tử protein là một trong những yếu tố ảnh hưởng đến độ hòa tan của chúng. Khi điện tích tại điểm đẳng điện bị mất đi, protein dễ dàng tổng hợp và kết tủa hơn. Điều này đặc biệt điển hình đối với các protein bị biến tính, trong đó các gốc axit amin kỵ nước xuất hiện trên bề mặt.
Trên bề mặt phân tử protein có cả gốc axit amin tích điện dương và âm. Số lượng các nhóm này và do đó tổng điện tích của protein phụ thuộc vào độ pH của môi trường, tức là. tỉ số nồng độ của nhóm H+ - và OH -. Trong môi trường axit Sự tăng nồng độ H+ dẫn đến ức chế sự phân ly của các nhóm carboxyl -COO - + H+ > - COOH và làm giảm điện tích âm của protein. Trong môi trường kiềm, sự liên kết của lượng dư OH - bởi proton hình thành trong quá trình phân ly nhóm amino -NH 3 + + OH - - NH 2 + H 2 O với sự hình thành nước, dẫn đến giảm điện tích dương của protein . Giá trị pH mà tại đó protein có điện tích bằng 0 được gọi là điểm đẳng điện (IEP). Trong IET, số lượng nhóm tích điện dương và âm là như nhau, tức là protein ở trạng thái đẳng điện.
3. Tách các protein riêng lẻ. Các đặc điểm về cấu trúc và chức năng của cơ thể phụ thuộc vào tập hợp protein được tổng hợp trong đó. Nghiên cứu cấu trúc và tính chất của protein là không thể nếu không tách chúng ra khỏi tế bào và tinh chế chúng khỏi các protein và phân tử hữu cơ khác. Các giai đoạn phân lập và tinh chế các protein riêng lẻ:
Phá hủy tế bào mô đang được nghiên cứu và thu được chất đồng nhất.
Tách chất đồng nhất thành các phân số bằng cách ly tâm, thu được phần nhân, ty thể, tế bào chất hoặc phần khác chứa protein mong muốn.
Biến tính nhiệt chọn lọc- đun nóng dung dịch protein trong thời gian ngắn, trong đó một số tạp chất protein bị biến tính có thể được loại bỏ (nếu protein tương đối ổn định nhiệt).
Muối ra. Các protein khác nhau kết tủa ở nồng độ muối khác nhau trong dung dịch. Bằng cách tăng dần nồng độ muối, có thể thu được một số phần riêng biệt với hàm lượng protein cô lập chiếm ưu thế trong một trong số chúng. Amoni sunfat thường được sử dụng để phân đoạn protein. Protein có độ hòa tan kém nhất sẽ kết tủa ở nồng độ muối thấp.
Lọc gel- phương pháp sàng lọc các phân tử qua hạt Sephadex trương nở (chuỗi polysaccharide ba chiều của dextran có lỗ xốp). Tốc độ protein đi qua cột chứa Sephadex sẽ phụ thuộc vào trọng lượng phân tử của chúng: khối lượng phân tử protein càng nhỏ thì chúng càng dễ thâm nhập vào các hạt và tồn tại ở đó lâu hơn; khối lượng càng lớn thì chúng rửa giải khỏi hạt càng nhanh. cột.
Siêu ly tâm- phương pháp liên quan đến việc đặt protein vào ống ly tâm vào rôto của máy siêu ly tâm. Khi rôto quay, tốc độ lắng của protein tỷ lệ thuận với trọng lượng phân tử của chúng: các phần protein nặng hơn nằm gần đáy ống nghiệm, các phần nhẹ hơn - gần bề mặt hơn.
Điện di- một phương pháp dựa trên sự khác biệt về tốc độ chuyển động của protein trong điện trường. Giá trị này tỷ lệ thuận với điện tích của protein. Điện di protein được thực hiện trên giấy (trong trường hợp này, tốc độ di chuyển của protein chỉ tỷ lệ thuận với điện tích của chúng) hoặc trong gel polyacrylamide có kích thước lỗ nhất định (tốc độ di chuyển của protein tỷ lệ thuận với điện tích và trọng lượng phân tử của chúng) .
Sắc kí trao đổi ion- phương pháp phân đoạn dựa trên sự liên kết của các nhóm protein bị ion hóa với các nhóm nhựa trao đổi ion tích điện trái dấu (vật liệu polyme không hòa tan). Độ bền liên kết của protein với nhựa tỷ lệ thuận với điện tích của protein. Các protein được hấp phụ vào polyme trao đổi ion có thể bị cuốn trôi khi tăng nồng độ dung dịch NaCl; Điện tích protein càng thấp thì nồng độ NaCl cần thiết để rửa trôi protein liên kết với các nhóm ion của nhựa càng thấp.
Sắc ký ái lực- phương pháp cụ thể nhất để tách riêng từng protein. Phối tử của protein được liên kết cộng hóa trị với một polyme trơ. Khi dung dịch protein được truyền qua cột có polyme, chỉ protein đặc hiệu cho phối tử nhất định mới được hấp phụ trên cột do liên kết bổ sung của protein với phối tử.
Chạy thận- phương pháp dùng để loại bỏ các hợp chất có trọng lượng phân tử thấp khỏi dung dịch protein đã phân lập. Phương pháp này dựa trên việc protein không thể đi qua màng bán thấm, không giống như các chất có trọng lượng phân tử thấp. Nó được sử dụng để tinh chế protein khỏi các tạp chất phân tử thấp, ví dụ như muối sau khi muối.
NHIỆM VỤ NGOẠI TRƯỢNG
1. Điền vào bảng. 1.4.
Bảng 1.4. Phân tích so sánh cấu trúc và chức năng của các protein liên quan - myoglobin và hemoglobin
a) nhớ cấu trúc trung tâm hoạt động của Mb và Hb. Các gốc axit amin kỵ nước đóng vai trò gì trong việc hình thành các trung tâm hoạt động của các protein này? Mô tả cấu trúc trung tâm hoạt động của Mb và Hb và cơ chế gắn kết của các phối tử với nó. Dư lượng His F 8 và His E 7 đóng vai trò gì trong hoạt động của trung tâm hoạt động của Mv iHv?
b) Protein oligomeric có liên quan chặt chẽ với nhau, hemoglobin, có những đặc tính mới nào so với myoglobin đơn phân? Giải thích vai trò của những thay đổi phối hợp trong cấu hình của các proton trong phân tử hemoglobin, ảnh hưởng của nồng độ CO 2 và proton đến ái lực của hemoglobin với oxy, cũng như vai trò của 2,3-BPG trong việc điều hòa dị lập thể của chức năng Hb .
2. Nêu đặc điểm của các chất đi kèm phân tử, chú ý đến mối quan hệ giữa cấu trúc và chức năng của chúng.
3. Những protein nào được nhóm lại thành các gia đình? Sử dụng ví dụ về họ globulin miễn dịch, hãy xác định các đặc điểm cấu trúc tương tự và chức năng liên quan của các protein thuộc họ này.
4. Các protein riêng lẻ đã được tinh chế thường được yêu cầu cho mục đích sinh hóa và y học. Giải thích các đặc tính hóa lý của protein mà các phương pháp được sử dụng để tách và tinh chế chúng dựa trên những đặc tính hóa lý nào.
NHIỆM VỤ TỰ KIỂM SOÁT
1. Chọn những đáp án đúng.
Chức năng của huyết sắc tố:
A. Vận chuyển O 2 từ phổi đến mô B. Vận chuyển H + từ mô đến phổi
B. Duy trì độ pH trong máu không đổi D. Vận chuyển CO 2 từ phổi đến các mô
D. Vận chuyển CO 2 từ mô đến phổi
2. Chọn những đáp án đúng. Phối tửα -protome Hb là: A. Heme
B. Ôxi
B. CO G. 2,3-BPG
D. β-Protom
3. Chọn những đáp án đúng.
Hemoglobin trái ngược với myoglobin:
A. Có cấu trúc bậc bốn
B. Cấu trúc bậc hai chỉ được biểu diễn bằng các chuỗi xoắn α
B. Thuộc loại protein phức tạp
D. Tương tác với phối tử dị lập thể D. Liên kết cộng hóa trị với heme
4. Chọn những đáp án đúng.
Ái lực của Hb với O2 giảm:
A. Khi thêm một phân tử O 2 B. Khi loại bỏ một phân tử O 2
B. Khi tương tác với 2,3-BPG
D. Khi gắn vào proton H + D. Khi nồng độ 2,3-BPG giảm
5. Cuộc thi đấu.
Các loại HB được đặc trưng bởi:
A. Ở dạng deoxy nó tạo thành các tập hợp fibrillar B. Chứa hai chuỗi α và hai chuỗi δ
B. Dạng Hb chiếm ưu thế trong hồng cầu trưởng thành D. Chứa heme và Fe+3 ở trung tâm hoạt động
D. Chứa hai chuỗi α và hai chuỗi γ 1. HbA 2.
6. Cuộc thi đấu.
Phối tử Hb:
A. Liên kết với Hb ở trung tâm dị lập thể
B. Có ái lực rất cao với trung tâm hoạt động của Hb
B. Bằng cách liên kết, nó làm tăng ái lực của Hb với O 2 G. Oxy hóa Fe+ 2 thành Fe+ 3
D. Hình thành liên kết cộng hóa trị với F8 của anh ấy
7. Chọn những đáp án đúng.
Người đi kèm:
A. Protein có ở mọi phần của tế bào
B. Sự tổng hợp tăng lên khi bị căng thẳng
B. Tham gia quá trình thủy phân protein bị biến tính
D. Tham gia duy trì cấu trúc tự nhiên của protein
D. Chúng tạo ra các bào quan trong đó hình thành cấu trúc của protein.
8. Trận đấu. Globulin miễn dịch:
A. Dạng bài tiết là ngũ sắc.
B. Lớp Ig xuyên qua hàng rào nhau thai
B. Ig - thụ thể tế bào mast
D. Lớp Ig chính có trong chất tiết của tế bào biểu mô. D. Thụ thể tế bào lympho B, sự kích hoạt của nó đảm bảo sự tăng sinh tế bào
9. Chọn những đáp án đúng.
Globulin miễn dịch E:
A. Được sản xuất bởi đại thực bào B. Chúng có chuỗi ε nặng.
B. Gắn vào màng tế bào lympho T
D. Đóng vai trò là thụ thể kháng nguyên màng trên tế bào mast và tế bào ưa bazơ
D. Chịu trách nhiệm về phản ứng dị ứng
10. Chọn những đáp án đúng.
Phương pháp tách protein dựa trên sự khác biệt về trọng lượng phân tử của chúng:
A. Lọc gel
B. Siêu ly tâm
B. Điện di trên gel polyacrylamide D. Sắc ký trao đổi ion
D. Sắc ký ái lực
11. Chọn câu trả lời đúng.
Phương pháp tách protein dựa trên sự khác biệt về độ hòa tan của chúng trong nước:
A. Lọc gel B. Muối ra
B. Sắc ký trao đổi ion D. Sắc ký ái lực
D. Điện di trên gel polyacrylamide
TIÊU CHUẨN ĐÁP ỨNG “NHIỆM VỤ TỰ KIỂM SOÁT”
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
ĐIỀU KHOẢN VÀ KHÁI NIỆM CƠ BẢN
1. Protein Oligomeric, protone, cấu trúc bậc bốn của protein
2. Những thay đổi hợp tác về hình dạng protone
3. Hiệu ứng Bohr
4. Điều hòa dị lập thể của các chức năng protein, trung tâm dị lập thể và tác nhân dị lập thể
5. Người đi kèm phân tử, protein sốc nhiệt
6. Họ protein (serine protease, immunoglobulin)
7. Mối quan hệ chức năng-cấu trúc IgM-, G-, E-, A
8. Tổng điện tích của protein, điểm đẳng điện của protein
9. Điện di
10. Đổ muối
11. Lọc gel
12. Sắc ký trao đổi ion
13. Siêu ly tâm
14. Sắc ký ái lực
15. Điện di protein huyết tương
NHIỆM VỤ CÔNG VIỆC TRONG LỚP
1. So sánh sự phụ thuộc của mức độ bão hòa của hemoglobin (Hb) và myoglobin (Mb) với oxy vào áp suất riêng phần của nó trong các mô
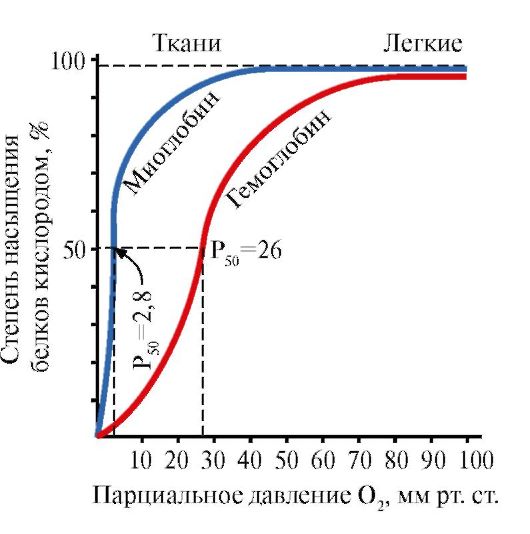 Cơm. 1,34. Sự phụ thuộc của độ bão hòa Mv vàNHoxy từ áp suất riêng phần của nó
Cơm. 1,34. Sự phụ thuộc của độ bão hòa Mv vàNHoxy từ áp suất riêng phần của nó
Xin lưu ý rằng hình dạng của đường cong bão hòa oxy trong protein là khác nhau: đối với myoglobin - hình hyperbol, đối với hemoglobin - hình sigmoid.
1. so sánh các giá trị áp suất riêng phần của oxy tại đó Mb và Hb bão hòa với O 2 bằng 50%. Protein nào sau đây có ái lực cao hơn với O 2?
2. Đặc điểm cấu trúc nào của Mb quyết định ái lực cao của nó với O 2?
3. Đặc điểm cấu trúc nào của HB cho phép nó giải phóng O2 trong các mao mạch của các mô đang nghỉ ngơi (ở áp suất riêng phần O2 tương đối cao) và làm tăng mạnh sự giải phóng này trong các cơ đang hoạt động? Tính chất nào của protein oligomeric mang lại hiệu ứng này?
4. Tính lượng O 2 (tính theo%) huyết sắc tố được oxy hóa cung cấp cho cơ bắp khi nghỉ ngơi và làm việc?
5. Rút ra kết luận về mối quan hệ giữa cấu trúc của protein và chức năng của nó.
2. Lượng oxy do hemoglobin giải phóng trong mao mạch phụ thuộc vào cường độ của quá trình dị hóa trong mô (hiệu ứng Bohr). Những thay đổi trong chuyển hóa mô điều chỉnh ái lực của Hb với O2 như thế nào? Ảnh hưởng của CO 2 và H+ đến ái lực của Hb với O 2
 1. mô tả hiệu ứng Bohr.
1. mô tả hiệu ứng Bohr.
2. quá trình thể hiện trong sơ đồ diễn ra theo hướng nào:
a) trong các mao mạch của phổi;
b) trong mao mạch mô?
3. Ý nghĩa sinh lý của hiệu ứng Bohr là gì?
4. Tại sao sự tương tác của Hb với H+ ở các vị trí xa heme lại làm thay đổi ái lực của protein đối với O 2?
3. Ái lực của Hb đối với O2 phụ thuộc vào nồng độ phối tử của nó - 2,3-bisphosphoglycerate, là chất điều chỉnh dị lập thể về ái lực của Hb đối với O2. Tại sao tương tác phối tử ở một vị trí cách xa vị trí hoạt động lại ảnh hưởng đến chức năng protein? 2,3-BPG điều chỉnh ái lực của Hb với O2 như thế nào? Để giải quyết vấn đề, hãy trả lời các câu hỏi sau:
1. 2,3-bisphosphoglycerate (2,3-BPG) được tổng hợp ở đâu và từ đâu? Viết công thức của nó, cho biết điện tích của phân tử này.
2. BPG tương tác với dạng hemoglobin (oxy hoặc deoxy) nào và tại sao? Sự tương tác xảy ra ở phần nào của phân tử Hb?
3. Quá trình thể hiện trong sơ đồ diễn ra theo hướng nào?
a) trong mao mạch mô;
b) trong mao mạch của phổi?
4. nơi có nồng độ phức chất cao hơn
Nv-2,3-BFG:
a) trong các mao mạch của cơ ở trạng thái nghỉ,
b) trong các mao mạch của cơ bắp đang hoạt động (được cung cấp cùng nồng độ BPG trong hồng cầu)?
5. Ái lực của HB đối với oxy sẽ thay đổi như thế nào khi con người thích nghi với điều kiện độ cao, nếu nồng độ BPG trong hồng cầu tăng lên? Ý nghĩa sinh lý của hiện tượng này là gì?
4. Sự phá hủy 2,3-BPG trong quá trình lưu trữ máu bảo quản sẽ làm suy yếu chức năng của HB. Ái lực của HB với O 2 trong máu bảo quản sẽ thay đổi như thế nào nếu nồng độ 2,3-BPG trong hồng cầu có thể giảm từ 8 xuống 0,5 mmol/l. Có thể truyền máu như vậy cho bệnh nhân bị bệnh nặng nếu nồng độ 2,3-BPG được phục hồi không sớm hơn sau ba ngày? Có thể khôi phục chức năng của hồng cầu bằng cách bổ sung 2,3-BPG vào máu không?
5. Hãy nhớ cấu trúc của các phân tử globulin miễn dịch đơn giản nhất. Globulin miễn dịch có vai trò gì trong hệ thống miễn dịch? Tại sao Igs thường được gọi là hóa trị hai? Cấu trúc của Igs liên quan đến chức năng của chúng như thế nào? (Mô tả bằng ví dụ về một nhóm globulin miễn dịch.)
Tính chất hóa lý của protein và phương pháp tách chúng.
6. Điện tích ròng của protein ảnh hưởng như thế nào đến độ hòa tan của nó?
a) xác định tổng điện tích của peptit ở pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) Điện tích của peptit này sẽ thay đổi như thế nào khi pH > 7, pH<7, рН <<7?
c) Điểm đẳng điện của protein (IEP) là gì và nó nằm trong môi trường nào?
IET của peptide này?
d) ở giá trị pH nào thì peptit này ít tan nhất.
7. Tại sao sữa chua, không giống như sữa tươi, lại “co lại” khi đun sôi (tức là protein sữa casein kết tủa)? Trong sữa tươi, phân tử casein mang điện tích âm.
8. Lọc gel được sử dụng để tách các protein riêng lẻ. Hỗn hợp chứa protein A, B, C có trọng lượng phân tử lần lượt là 160.000, 80.000 và 60.000 được phân tích bằng phương pháp lọc gel (Hình 1.35). Các hạt gel trương nở có khả năng thấm vào các protein có trọng lượng phân tử nhỏ hơn 70.000. Nguyên lý nào làm nền tảng cho phương pháp tách này? Đồ thị nào phản ánh chính xác kết quả phân số? Cho biết thứ tự các protein A, B, C được giải phóng khỏi cột.
 Cơm. 1,35. Sử dụng phương pháp lọc gel để tách protein
Cơm. 1,35. Sử dụng phương pháp lọc gel để tách protein
9. Trong bộ lễ phục. Hình 1.36, A thể hiện sơ đồ điện di trên giấy protein huyết thanh của người khỏe mạnh. Lượng tương đối của các phần protein thu được bằng phương pháp này là: albumin 54-58%, α 1 -globulin 6-7%, α 2 -globulin 8-9%, β-globulin 13%, γ-globulin 11-12%.
 Cơm. 1.36 Điện di trên giấy protein huyết tương của người khỏe mạnh (A) và bệnh nhân (B)
Cơm. 1.36 Điện di trên giấy protein huyết tương của người khỏe mạnh (A) và bệnh nhân (B)
I - γ-globulin; II - β-globulin; III -α 2 -globulin; IV -α 2 -globulin; V - albumin
Nhiều bệnh đi kèm với những thay đổi về số lượng trong thành phần protein huyết thanh (rối loạn protein máu). Bản chất của những thay đổi này được tính đến khi đưa ra chẩn đoán và đánh giá mức độ nghiêm trọng cũng như giai đoạn của bệnh.
Sử dụng số liệu cho trong bảng. 1.5, hãy đoán về căn bệnh này, được đặc trưng bởi hồ sơ điện di được trình bày trong Hình. 1,36.
Bảng 1.5. Sự thay đổi nồng độ protein huyết thanh trong bệnh lý
Trung tâm hoạt động của protein làđây là vị trí liên kết protein-phối tử. Một vùng được hình thành trên bề mặt của hạt có thể gắn các phân tử khác gọi là phối tử . Trung tâm hoạt động của protein được hình thành từ các nhóm axit amin bên được tập hợp lại với nhau ở cấp độ cấu trúc bậc ba. Trong trình tự tuyến tính của chuỗi peptide, chúng có thể nằm ở khoảng cách rất xa nhau. Protein thể hiện tính đặc hiệu cao khi tương tác với phối tử. Độ đặc hiệu cao của sự tương tác của protein với phối tử được đảm bảo bởi tính bổ sung giữa cấu trúc của trung tâm hoạt động của protein với cấu trúc của phối tử. Tính bổ sung – đây là sự tương ứng về không gian và hóa học của các phân tử tương tác. Các trung tâm liên kết protein-phối tử thường nằm giữa các miền (ví dụ trung tâm liên kết của trypsin với phối tử của nó có 2 miền cách nhau bằng một rãnh).
Chức năng của protein dựa trên sự tương tác cụ thể của chúng với các phối tử. 50.000 protein riêng lẻ chứa các trung tâm hoạt động duy nhất chỉ có thể liên kết với các phối tử cụ thể và do đặc điểm cấu trúc của trung tâm hoạt động, thể hiện các chức năng vốn có của chúng. Rõ ràng, cấu trúc bậc một chứa thông tin về chức năng của protein.
Cấu trúc bậc bốn- đây là cấp độ tổ chức cấu trúc cao nhất, không phải loại protein nào cũng có được. Cấu trúc bậc bốn được hiểu là cách các chuỗi polypeptide được sắp xếp trong không gian và hình thành một quần thể cao phân tử thống nhất về mặt cấu trúc và chức năng. Mỗi chuỗi polypeptide riêng biệt được gọi là proton hoặc tiểu đơn vị , thường không có hoạt tính sinh học. Protein có được khả năng này thông qua một phương pháp liên kết không gian nhất định của các proton cấu thành của nó. Phân tử thu được thường được gọi là oligome (đa bội) .
Cấu trúc bậc bốn được ổn định nhờ các liên kết không cộng hóa trị phát sinh giữa các vùng tiếp xúc của proton, chúng tương tác với nhau theo kiểu bổ sung.
Protein có cấu trúc bậc bốn bao gồm nhiều enzyme (lactate dehydrogenase, glutamate dehydrogenase, v.v.), cũng như hemoglobin, protein myosin của cơ co bóp. Một số protein có số lượng tiểu đơn vị nhỏ, từ 2 đến 8, trong khi những protein khác có hàng trăm, thậm chí hàng nghìn tiểu đơn vị. Ví dụ, protein virus khảm thuốc lá có 2130 tiểu đơn vị.
Một ví dụ điển hình của protein có cấu trúc bậc bốn là hemoglobin. Phân tử hemoglobin gồm 4 tiểu đơn vị là chuỗi polypeptide, mỗi chuỗi liên kết với heme, trong đó có 2 chuỗi polypeptide là -2afla và -2beta, chúng khác nhau về cấu trúc bậc một và độ dài của chuỗi polypeptide.
Các liên kết hình thành cấu trúc bậc bốn kém bền hơn. Dưới tác dụng của một số tác nhân, protein được chia thành các tiểu đơn vị riêng biệt. Khi tác nhân bị loại bỏ, các tiểu đơn vị có thể kết hợp lại và chức năng sinh học của protein được phục hồi. Vì vậy, khi urê được thêm vào dung dịch huyết sắc tố, nó sẽ phân hủy thành 4 tiểu đơn vị cấu thành; khi loại bỏ urê, vai trò cấu trúc và chức năng của huyết sắc tố được phục hồi.
Kết thúc công việc -
Chủ đề này thuộc chuyên mục:
Hóa sinh. Sóc. Axit amin là thành phần cấu trúc của protein
Protein axit amin thành phần cấu trúc của protein.. protein.. protein là các hợp chất hữu cơ phân tử cao chứa nitơ bao gồm các axit amin nối thành chuỗi với ..
Nếu bạn cần thêm tài liệu về chủ đề này hoặc bạn không tìm thấy những gì bạn đang tìm kiếm, chúng tôi khuyên bạn nên sử dụng tìm kiếm trong cơ sở dữ liệu tác phẩm của chúng tôi:
Chúng ta sẽ làm gì với tài liệu nhận được:
Nếu tài liệu này hữu ích với bạn, bạn có thể lưu nó vào trang của mình trên mạng xã hội:
| tiếng riu ríu |
Tất cả các chủ đề trong phần này:
Cơ chế hoạt động của enzyme
Theo các khái niệm hiện đại, sự tương tác của enzyme với cơ chất có thể được chia thành 3 giai đoạn: Giai đoạn 1 được đặc trưng bởi sự khuếch tán của cơ chất đến enzyme.
Xúc tác axit-bazơ
Vị trí hoạt động của enzyme chứa các nhóm axit và cơ bản. Các nhóm loại axit loại bỏ H+ và mang điện tích âm. Các nhóm thuộc loại cơ bản gắn H+ và có một nửa
MỘT). Giả thuyết của Fisher
Theo đó, có sự tương ứng không gian chặt chẽ giữa cơ chất và trung tâm hoạt động của enzyme. Theo Fischer, enzyme là một cấu trúc cứng nhắc và cơ chất là một khối của trung tâm hoạt động của nó.
Sự trao đổi carbohydrate
CHUYỂN HÓA CARBOHYDRATE 1. Carbohydrate chính của cơ thể động vật, vai trò sinh học của chúng. 2. Chuyển hóa carbohydrate trong các cơ quan của hệ tiêu hóa. 3. Sinh tổng hợp và thoái hóa
Vai trò sinh học của carbohydrate
VAI TRÒ SINH HỌC CỦA CARBOHYDRATE: 1. NĂNG LƯỢNG. Khi 1 g carbohydrate bị oxy hóa thành sản phẩm cuối cùng (CO2 và H2O), 4,1 kcal năng lượng được giải phóng. Tỷ lệ carbohydrate là khoảng 60-70
Chuyển hóa carbohydrate trong đường tiêu hóa
CHUYỂN ĐỔI CARBOHYDRATE TRONG ĐƯỜNG TIÊU HÓA Carbohydrate chính trong thức ăn cho cơ thể con người là: tinh bột, glycogen, sucrose, lactose. Tinh bột hấp thụ từ thức ăn
Sinh tổng hợp và phân hủy glycogen
SINH TỔNG HỢP VÀ HÒA HÒA GLYCOGEN TRONG MÔ. BỆNH GLYCOGEN. Người ta phát hiện ra rằng glycogen có thể được tổng hợp ở hầu hết các cơ quan và mô. Tuy nhiên, kết thúc lớn nhất của nó
Glycolysis kỵ khí
Tùy thuộc vào trạng thái chức năng của cơ thể, các tế bào của các cơ quan và mô có thể ở trong tình trạng được cung cấp đủ oxy hoặc bị thiếu oxy, sau đó
Đường phân hiếu khí (con đường hexose diphosphate)
CON ĐƯỜNG HEXOSODIPHOSPHATE. Đây là con đường cổ điển của quá trình dị hóa hiếu khí carbohydrate trong mô, diễn ra trong tế bào chất cho đến giai đoạn hình thành pyruvate và kết thúc ở ty thể với sự hình thành các hợp chất.
Con đường hexose monophosphate
CON ĐƯỜNG HEXOSMONOPHOSPHATE ĐỂ CHUYỂN ĐỔI GLUCOSE TRONG MÔ, PHẢN ỨNG HÓA HỌC. Quá trình oxy hóa glucose dọc theo con đường này xảy ra trong tế bào chất của tế bào và được thể hiện bằng hai nhánh liên tiếp
Tân tạo glucose
TỔNG HỢP GLUCON Các nguồn cung cấp glucose chính cho cơ thể con người là: 1. carbohydrate trong chế độ ăn uống; 2. glycogen mô; 3. tân tạo đường. GLUCONOEGENESIS là
Các lipid chính của cơ thể con người, vai trò sinh học của chúng
LIPIDS là các chất hữu cơ phức tạp có bản chất sinh học, không hòa tan trong nước nhưng hòa tan trong dung môi hữu cơ. LIPIDS là một loại thực phẩm chủ yếu. họ đang
Tiêu hóa lipid, tái tổng hợp chất béo
Tiêu hóa lipid. LIPIDS đến từ thức ăn trong khoang miệng chỉ được xử lý cơ học. Enzym LIPOLYTIC không được sản xuất trong khoang miệng. Tiêu hóa chất béo
Lipoprotein máu
LIPIDS là những hợp chất không tan trong nước nên việc vận chuyển chúng trong máu cần có chất mang đặc biệt hòa tan trong nước. Các hình thức vận chuyển như vậy là LIPOPROTEIN.
Quá trình oxy hóa các axit béo cao hơn
Mô mỡ, bao gồm các tế bào mỡ, đóng một vai trò cụ thể trong chuyển hóa lipid. Khoảng 65% khối lượng mô mỡ đến từ triacylglycerol (TAG) tích tụ trong đó - chúng đại diện cho
Sinh tổng hợp IVF trong mô
Sinh tổng hợp IVH xảy ra trong mạng lưới nội chất của tế bào. IVA có thể thay thế (tất cả đều bão hòa và không bão hòa, có một liên kết đôi) được tổng hợp trong các tế bào từ ACETYL-CoA. Điều kiện để có bi
Trao đổi cholesterol
Trao đổi cholesterol. Cholesterol là tiền chất trong quá trình tổng hợp các steroid: acid mật, hormone steroid, vitamin D3. Cholesterol là thành phần cấu trúc bắt buộc
Tiêu hóa protein
Tiêu hóa protein trong đường tiêu hóa Protein thực phẩm trải qua quá trình thủy phân dưới tác dụng của ENZYME PROTEOLYTIC (lớp - hydrolase, phân lớp - peptidase).
Quá trình thối rữa axit amin, trung hòa sản phẩm thối rữa
RỘNG RÃI CỦA AXIT AMIN Axit amin không được hấp thụ sẽ đi vào ruột già, nơi chúng bị thối rữa. QUAY QUAY AXIT AMIN là quá trình phân hủy các axit amin dưới tác dụng của
Chuyển hóa axit amin
Chuyển hóa axit amin Nguồn cung cấp axit amin trong tế bào là: 1. Protein thực phẩm sau khi thủy phân ở cơ quan tiêu hóa; 2. tổng hợp các axit amin không thiết yếu;
Các cách trung hòa amoniac
Amoniac được hình thành từ các axit amin trong quá trình phân hủy các hợp chất chứa nitơ khác (amine sinh học, NUCLEOTIDES). Một phần đáng kể amoniac được hình thành trong ruột già trong quá trình thối rữa. Nó bị hút vào
Điều hòa trao đổi chất
PHÂN TÍCH TÍN HIỆU. Nhiệm vụ chính của việc điều hòa quá trình trao đổi chất và chức năng tế bào: 1. Sự phối hợp nội bào và giữa các quá trình trao đổi chất; 2. loại trừ “nhàn rỗi”
Hormon vùng dưới đồi
HYPOTHALAMUS HORMONES HYPOTHALAMUS là một thành phần và một loại “kênh đầu ra” của hệ thống limbic. Đây là một phần của gian não kiểm soát các thông số khác nhau của não
Hormon tuyến yên
Hormon tuyến yên
HORMON TUYẾN TUYẾN Tuyến yên được chia thành thùy trước (adenohypophys) và thùy sau (neurohypophys). Các hormone của tuyến yên có thể được chia thành 3 nhóm tùy thuộc vào
Sinh tổng hợp iodothyronine
Sự tổng hợp iodothyronine xảy ra như một phần của protein - thyroglobulin, nằm trong các nang của tuyến giáp. Thyroglobulin là một glycoprotein chứa 115 gốc tyrosine. P
Chuyển hóa lipid
Trong mô mỡ gan, hormone kích thích quá trình phân giải lipid. Những tác động này lên quá trình chuyển hóa carbohydrate và lipid có liên quan đến sự gia tăng độ nhạy cảm của tế bào đối với hoạt động của adrenaline dưới tác động của hormone tuyến giáp.
giảm tiết
Ở thời thơ ấu, sự bài tiết giảm dẫn đến chậm phát triển về thể chất và tinh thần (đờ đẫn). Ở người lớn, biểu hiện nặng của tình trạng thiếu hụt hormone tuyến giáp được trộn lẫn
tăng tiết
Bướu cổ nhiễm độc lan tỏa (bệnh Graves) là bệnh phổ biến nhất kèm theo tăng sản xuất iodothyronine. Với căn bệnh này, kích thước của tuyến giáp tăng lên và
Hormon tuyến cận giáp
Hormon tuyến cận giáp được tổng hợp ở tuyến cận giáp và bao gồm 84 dư lượng axit amin. Hormon được lưu trữ trong các hạt bài tiết. Sự tiết PTH được điều hòa bởi nồng độ canxi trong máu: trong khi ngủ
Hormon tuyến sinh dục
Hormon tuyến sinh dục là steroid có bản chất hóa học. Có: 1. Androgen; 2. Nội tiết tố nữ; 3. Progestin.
Hormon tuyến thượng thận
Hormon tuyến thượng thận Tuyến thượng thận là tuyến nội tiết tiết ra vỏ não và tủy. Các hormone steroid được tổng hợp ở vỏ não và trong não
Hormon tuyến tụy
Các hormone của tuyến tụy Chức năng của tuyến tụy: · ngoại tiết; · Nội tiết. Chức năng ngoại tiết là tổng hợp và bài tiết chất tiêu hóa
Câu hỏi thi
KHOA DƯỢC PHẨM (Khoa PHƯƠNG PHÁP) Đề thi môn Hóa sinh cho sinh viên năm thứ 3 (học kỳ 6) 1. Hóa sinh, nhiệm vụ của nó. Mối liên hệ giữa sinh hóa và f
CẤU HÌNH VÀ CẤU HÌNH CỦA PHÂN TỬ PROTEIN
Kính hiển vi điện tử
Có thể dùng để xác định cấu trúc của các phân tử protein có trọng lượng phân tử lớn - từ 500.000 đến 1.000.000 Da (dalton). Dalton (Da) và kilodalton (kDa)– đơn vị đo khối lượng protein. 1kDa=10 3 Có. 1 dalton bằng 1/16 khối lượng của một nguyên tử oxy (đơn vị khối lượng oxy).
Từ tất cả những gì đã nói, chúng ta có thể kết luận rằng tổ chức không gian của protein rất phức tạp. Trong hóa học có một khái niệm - không gian CẤU HÌNH - Sự sắp xếp tương đối về mặt không gian của các phần của phân tử được cố định chặt chẽ bằng liên kết cộng hóa trị(ví dụ: thuộc dòng L của các đồng phân lập thể hoặc thuộc dòng D).
Đối với protein, khái niệm này cũng được sử dụng CẤU HÌNH phân tử protein - sự sắp xếp tương đối xác định, nhưng không cố định, không thể thay đổi của các bộ phận của phân tử. Vì cấu trúc của phân tử protein được hình thành với sự tham gia của các loại liên kết yếu nên nó có tính di động (có khả năng thay đổi) và protein có thể thay đổi cấu trúc của nó. Tùy thuộc vào điều kiện môi trường, một phân tử có thể tồn tại ở các trạng thái hình dạng khác nhau, dễ dàng biến đổi lẫn nhau. Thuận lợi về mặt năng lượng đối với các điều kiện thực tế chỉ là một hoặc một số trạng thái hình dạng mà trong đó có sự cân bằng. Sự chuyển đổi từ trạng thái hình dạng này sang trạng thái hình dạng khác đảm bảo hoạt động của phân tử protein. Đây là những thay đổi về hình dạng có thể đảo ngược (ví dụ được tìm thấy trong cơ thể trong quá trình dẫn truyền xung thần kinh, trong quá trình vận chuyển oxy bằng huyết sắc tố). Khi hình dạng thay đổi, một số liên kết yếu bị phá hủy và các liên kết yếu mới được hình thành.
Sự tương tác của protein với một chất đôi khi dẫn đến sự liên kết giữa một phân tử của chất này với một phân tử protein. Hiện tượng này được gọi là "hấp thụ" (ràng buộc). Quá trình ngược lại - giải phóng một phân tử khác từ protein được gọi là "giải hấp".
Nếu đối với một số cặp phân tử, quá trình hấp phụ chiếm ưu thế so với quá trình giải hấp thì điều này đã sự hấp thụ cụ thể, và chất bị hấp phụ gọi là "phối tử".
Các loại phối tử:
1) Phối tử của protein enzyme là cơ chất.
2) Phối tử protein vận chuyển – chất được vận chuyển.
3) Phối tử kháng thể (immunoglobulin) – kháng nguyên.
4) Phối tử thụ thể hormone hoặc chất dẫn truyền thần kinh - hormone hoặc chất dẫn truyền thần kinh.
Protein có thể thay đổi cấu hình của nó không chỉ khi tương tác với phối tử mà còn do bất kỳ tương tác hóa học nào. Một ví dụ về sự tương tác như vậy là việc bổ sung dư lượng axit photphoric.
Trong điều kiện tự nhiên, protein có một số trạng thái hình dạng thuận lợi về mặt nhiệt động. Đây là những trạng thái bản địa (tự nhiên). Natura (lat.) - thiên nhiên.


